Cas9 vs Cas12a for Multiplex Editing: A Strategic Guide for Therapeutic Development
This article provides a comprehensive comparison of CRISPR-Cas9 and Cas12a systems, with a dedicated focus on their application in multiplex genome editing for biomedical research and therapeutic development.
Cas9 vs Cas12a for Multiplex Editing: A Strategic Guide for Therapeutic Development
Abstract
This article provides a comprehensive comparison of CRISPR-Cas9 and Cas12a systems, with a dedicated focus on their application in multiplex genome editing for biomedical research and therapeutic development. It covers foundational mechanisms, PAM requirements, and inherent multiplex capabilities. The content explores advanced methodologies for immune cell engineering and in vivo disease modeling, addresses critical troubleshooting aspects like editing efficiency and off-target effects, and presents validation data from recent comparative studies. Aimed at researchers and drug development professionals, this guide synthesizes current evidence to inform strategic tool selection for complex genetic perturbations.
CRISPR Cas9 and Cas12a: Unpacking Core Mechanisms for Multiplex Editing
Distinct Evolutionary Origins and Molecular Architectures of Cas9 and Cas12a
The discovery of CRISPR-Cas systems has revolutionized genome editing, yet the distinct properties of different Cas nucleases determine their suitability for specific research applications. Among these, Cas9 and Cas12a represent two fundamentally different classes of RNA-guided nucleases with unique evolutionary backgrounds and structural architectures [1]. While Cas9 from Streptococcus pyogenes became the pioneering tool for genome engineering, Cas12a (formerly Cpf1) emerged as a powerful alternative with contrasting molecular mechanisms that influence its performance in multiplex editing, target specificity, and editing outcomes [2]. Understanding these differences is crucial for researchers selecting the optimal system for genetic screening, therapeutic development, and functional genomics. This guide provides a comprehensive comparison of these systems, supported by experimental data and protocol details, to inform their application in multiplex editing research.
Fundamental Structural and Functional Divergence
Cas9 and Cas12a originate from distinct evolutionary pathways within bacterial immune systems, resulting in significant structural differences that dictate their molecular functions [1].
Molecular Architecture and DNA Cleavage Mechanisms
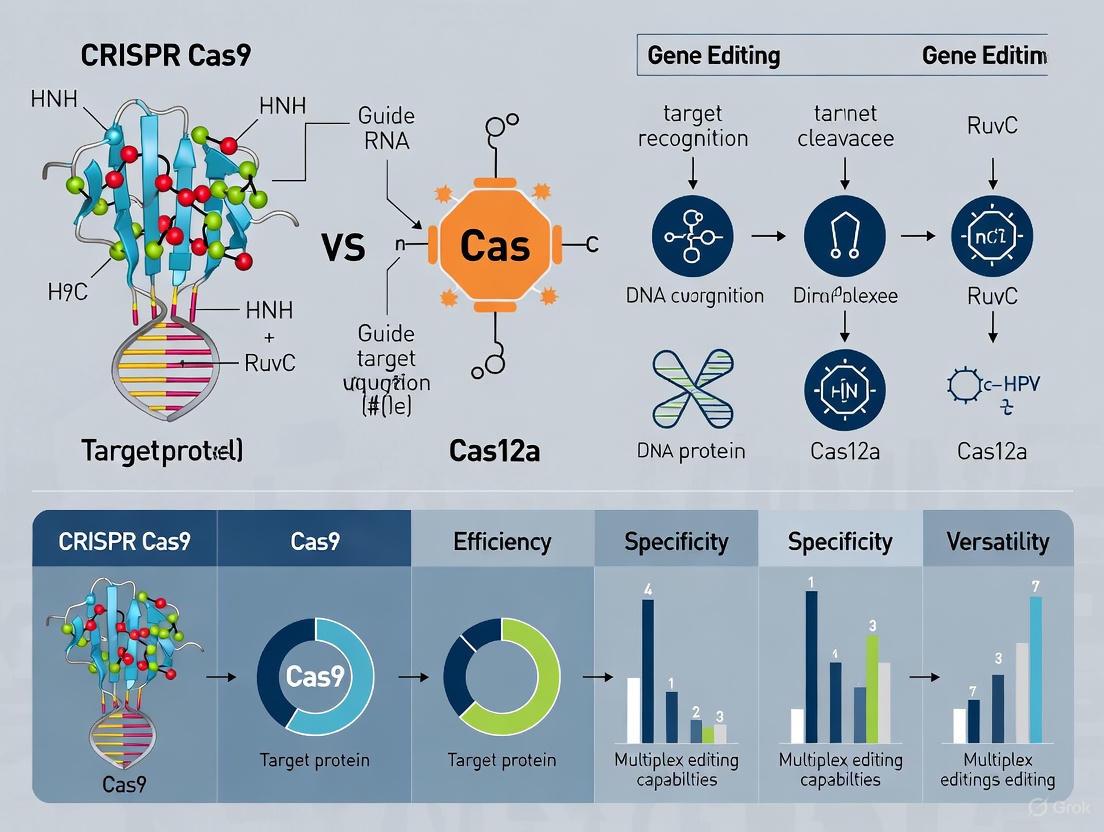
Guide RNA Requirements: Cas9 necessitates a dual RNA system, typically combined into a single guide RNA (sgRNA), comprising both CRISPR RNA (crRNA) and trans-activating crRNA (tracrRNA) components. In contrast, Cas12a requires only a single crRNA for DNA recognition and cleavage, simplifying guide design and reducing the payload size [3].
DNA Cleavage Characteristics: Cas9 generates blunt-ended double-strand breaks 3-4 base pairs upstream of the protospacer adjacent motif (PAM). Cas12a creates staggered cuts with 5' overhangs, which can facilitate more predictable repair outcomes and potentially enhance homology-directed repair efficiency [2].
PAM Recognition Specificity: Cas9 recognizes G-rich PAM sequences (5'-NGG-3'), while Cas12a binds to T-rich PAM sequences (5'-TTTV-3'), significantly expanding targeting capabilities in AT-rich genomic regions that are challenging for Cas9 [2].
Table 1: Fundamental Characteristics of Cas9 and Cas12a
| Feature | Cas9 | Cas12a |
|---|---|---|
| Evolutionary Origin | Type II CRISPR system | Type V-A CRISPR system |
| Guide RNA | Dual RNA (crRNA + tracrRNA) or sgRNA | Single crRNA |
| crRNA Length | ~100 nt for sgRNA | ~42-44 nt |
| PAM Sequence | 5'-NGG-3' (G-rich) | 5'-TTTV-3' (T-rich) |
| DNA Cleavage | Blunt ends | Staggered cuts with 5' overhangs |
| RNase Activity | No | Yes (processes its own crRNA arrays) |
Quantitative Performance Comparison in Genome Editing
Recent comparative studies directly assess the editing profiles and efficiencies of Cas9 and Cas12a across various biological systems, providing crucial data for experimental planning.
Editing Efficiency and Mutation Profiles
A comprehensive 2024 study in tomato protoplasts systematically compared LbCas12a and SpCas9 editing efficiencies at 35 overlapping target sites [4]. The research demonstrated that while both nucleases achieved effective mutagenesis, Cas12a induced more and larger deletions than Cas9, a characteristic potentially advantageous for gene knockout applications. The study found similar overall editing efficiencies between the two systems, though performance was target-dependent [4].
Table 2: Experimental Editing Outcomes in Tomato Protoplasts [4]
| Parameter | SpCas9 | LbCas12a |
|---|---|---|
| Average Mutation Efficiency | Target-dependent, high | Target-dependent, comparable to Cas9 |
| Primary Mutation Type | Small insertions/deletions | Larger deletions |
| Deletion Size Range | Typically 1-10 bp | Often exceeds 10 bp |
| Off-target Rate | Lower at sites with >2 mismatches | 10/57 predicted off-target sites edited |
| Mismatch Tolerance | Higher, especially distal to PAM | Strict requirement for perfect base pairing |
Specificity and Off-Target Considerations
Thermodynamic studies reveal fundamental differences in how Cas9 and Cas12a tolerate mismatches between guide RNA and target DNA. Research indicates that Cas12a-crRNA complexes demonstrate greater thermodynamic stability with perfectly matched DNA but experience more significant destabilization from mismatches compared to Cas9-sgRNA complexes [5]. This characteristic translates to reduced off-target activity for Cas12a in many genomic contexts, though both systems require careful guide design to minimize off-target effects [4].
Analysis of off-target editing in tomato cells revealed that all identified Cas12a off-target sites contained only one or two mismatches distal from the PAM, highlighting the importance of comprehensive off-target prediction in experimental design [4].
Multiplex Editing Capabilities and crRNA Processing
A significant advantage of Cas12a for complex genome engineering applications lies in its intrinsic capacity for multiplexed editing through coordinated processing of crRNA arrays.
Native Multiplexing Through crRNA Processing
Unlike Cas9, which requires additional RNA-processing systems (tRNA, ribozymes, or Csy4) for multiplexing, Cas12a possesses intrinsic RNase activity that enables it to process a single transcript containing multiple crRNAs into individual functional guides [6]. This native capability simplifies vector design for multiplexed editing applications, as researchers can deliver a single array encoding multiple guides rather than constructing complex multi-promoter systems [6].
The molecular basis for this function stems from Cas12a's ability to recognize direct repeats flanking crRNA sequences in a long transcript and cleave them into mature crRNAs, enabling simultaneous targeting of multiple genomic loci from a single expression cassette [6].
Experimental Workflow for Cas12a Multiplex Editing
The typical workflow for implementing Cas12a multiplex editing involves:
- Array Design: Designing a crRNA array with multiple targeting sequences separated by direct repeats
- Vector Construction: Cloning the array into an appropriate expression vector
- Delivery: Introducing the Cas12a nuclease and crRNA array into target cells
- Processing: Endogenous cleavage of the array by Cas12a into individual crRNAs
- Editing: Simultaneous targeting of multiple genomic loci
This streamlined approach has been successfully implemented in combinatorial genetic screening in mammalian cells, demonstrating the practical utility of Cas12a for complex editing applications [6].
Engineering and Optimization Strategies
Both Cas9 and Cas12a have undergone extensive protein engineering to enhance their functionality and address limitations in native systems.
Enhanced Cas12a Variants for Improved Performance
Initial implementations of Cas12a in mammalian cells showed lower editing efficiency compared to Cas9, prompting engineering efforts to improve performance [6]. Key optimization strategies include:
Nuclear Localization Enhancement: Addition of multiple nuclear localization signals (NLS) to improve nuclear import, with studies showing 4- to 32-fold improvement with six NLS additions [6]
crRNA Engineering: Incorporating a direct repeat sequence 3' of the crRNA to enhance stability and editing efficiency, approximately doubling knockout efficiency [6]
Protein Mutations: Introducing point mutations (E174R, S542R for AsCas12a) to increase DNA binding affinity and cleavage activity [6]
These combined optimizations have yielded variants such as "opAsCas12a" (optimized AsCas12a) with 32- to 64-fold improvement over baseline Cas12a performance, making it competitive with SpCas9 in dropout screening applications [6].
PAM Expansion and Specificity Engineering
Recent directed evolution approaches have addressed Cas12a's restricted PAM requirements (5'-TTTV-3'), which limited targetable sites to approximately 1% of the genome [7]. Through bacterial selection systems and rational engineering, researchers developed Flex-Cas12a variants with expanded PAM recognition (5'-NYHV-3'), increasing potential target sites to over 25% of the human genome while maintaining recognition of canonical PAMs [7].
Similar engineering efforts have created high-fidelity Cas9 variants with reduced off-target effects, highlighting how both systems continue to evolve through protein engineering to overcome inherent limitations.
Table 3: Engineered Variants and Their Applications
| Variant | Key Modifications | Performance Improvements | Primary Applications |
|---|---|---|---|
| opAsCas12a | 6xNLS, dual-DR crRNA, E174R/S542R mutations | 32-64x efficiency improvement | Combinatorial genetic screening [6] |
| hyperCas12a | D156R/D235R/E292R/D350R mutations | Enhanced activity at low crRNA concentrations | In vivo multiplexed gene regulation [8] |
| Flex-Cas12a | G146R/R182V/D535G/S551F/D665N/E795Q | Recognizes 5'-NYHV-3' PAMs (~25% genome coverage) | Access to previously inaccessible loci [7] |
| enAsCas12a | E174R/S542R/K548R mutations | Increased DNA binding affinity | General genome editing applications [6] |
Experimental Design and Protocol Considerations
Research Reagent Solutions
Table 4: Essential Reagents for Cas12a-based Genome Editing
| Reagent | Function | Examples & Specifications |
|---|---|---|
| Cas12a Nuclease | RNA-guided DNA endonuclease | LbCas12a, AsCas12a, FnCas12a (codon-optimized for expression system) |
| crRNA Expression Vector | Guide RNA delivery | U6 promoter-driven crRNA with appropriate direct repeats |
| crRNA Array | Multiplex editing | Multiple crRNAs separated by direct repeats in single transcript |
| Delivery System | Cellular introduction of editing components | Lentivirus, AAV, lipid nanoparticles (LNPs), electroporation |
| NLS Tags | Nuclear localization | SV40, c-Myc, or bipartite NLS (multiple copies enhance efficiency) |
| Selection Marker | Transformed cell enrichment | Antibiotic resistance, fluorescent proteins (GFP, RFP) |
Detailed Methodology for Cas12a Multiplex Editing
Based on established protocols from recent studies [6], the following workflow enables efficient Cas12a multiplex editing:
Vector Construction Protocol:
- crRNA Array Design: Design 2-4 crRNA sequences targeting desired genomic loci, each flanked by the appropriate direct repeat sequences (varies by Cas12a ortholog)
- Golden Gate Assembly: Clone synthesized oligos into BsaI-digested crRNA expression vector using MoClo toolkit components [4]
- Nuclease Vector Preparation: Clone codon-optimized Cas12a with N-terminal and C-terminal NLS tags into mammalian expression vector under EF1α or similar promoter
- Validation: Sanger sequence final constructs to verify correct assembly
Cell Culture and Transfection:
- Cell Line Preparation: Culture appropriate cell line (e.g., K562, HEK293T, or target-specific lines) to 70-80% confluence
- Lentiviral Production: For stable expression, produce lentivirus in HEK293T cells using psPAX2 and pMD2.G packaging plasmids
- Transduction: Transduce target cells at appropriate MOI, then select with puromycin (or relevant antibiotic) for 5-7 days
- Editing Validation: Harvest genomic DNA 7-14 days post-transduction; assess editing efficiency via T7E1 assay or next-generation amplicon sequencing
Experimental Validation for Multiplex Editing:
- Amplicon Sequencing: Design primers flanking each target site; perform PCR and NGS library preparation
- Data Analysis: Use computational pipelines (CRISPResso2, etc.) to quantify insertion/deletion frequencies at each target
- Off-target Assessment: Perform Cas-OFFinder prediction [4] and examine top potential off-target sites via amplicon sequencing
The distinct evolutionary origins and molecular architectures of Cas9 and Cas12a have produced complementary genome editing systems with unique strengths. Cas12a offers significant advantages for multiplex editing applications due to its intrinsic crRNA processing capabilities, staggered cut formation, and stringent mismatch sensitivity [6]. The development of high-performance variants like opAsCas12a and hyperCas12a has addressed initial efficiency concerns, making Cas12a competitive with Cas9 while retaining its unique functional benefits [6] [8].
For researchers designing multiplex editing experiments, Cas12a provides a streamlined solution for simultaneous targeting of multiple genomic loci without requiring complex multi-guide expression systems. Its preference for T-rich PAM sequences additionally enables targeting of genomic regions inaccessible to Cas9 [2]. Continued protein engineering efforts are further expanding the targeting range and specificity of both systems, providing researchers with an increasingly sophisticated toolkit for precise genome manipulation [7]. The strategic selection between Cas9 and Cas12a should be guided by specific experimental requirements including target sequence context, desired editing outcomes, multiplexing scale, and specificity constraints.
In the realm of CRISPR-based genome editing, the Protospacer Adjacent Motif (PAM) serves as the essential gateway that dictates a nuclease's access to the genome. This short, specific DNA sequence flanking the target site is not merely a binding preference but a fundamental requirement for initiating the editing process. For researchers, scientists, and drug development professionals engaged in multiplex editing research, the choice between CRISPR-Cas9 and CRISPR-Cas12a often hinges on their distinct PAM requirements: the G-rich NGG for Cas9 versus the T-rich TTTV for Cas12a. This comparison guide objectively analyzes how these PAM specificities shape targetable genomic space, editing efficiency, and experimental outcomes, providing a scientific foundation for strategic nuclease selection in complex genome engineering projects. Understanding these differences is crucial for expanding the accessible genome and improving the precision of genetic modifications across diverse applications from basic research to therapeutic development.
Fundamental Differences in PAM Recognition
The PAM sequences recognized by Cas9 and Cas12a represent one of the most distinguishing features between these two CRISPR systems. Cas9 from Streptococcus pyogenes (SpCas9), the most widely utilized variant, requires a 5'-NGG-3' PAM sequence located immediately downstream of the target DNA sequence [9]. This PAM is relatively abundant in many genomes but presents limitations in AT-rich regions where G-content is sparse. In contrast, Cas12a (formerly known as Cpf1) recognizes a 5'-TTTV-3' PAM (where V is A, C, or G) located upstream of the target sequence [10] [2]. This fundamental difference in PAM recognition orientation and sequence preference directly influences the targetable regions within any given genome.
The structural biology underlying PAM recognition further differentiates these systems. Cas9 employs a PAM-interacting domain within its recognition lobe to engage with the G-rich PAM sequence, while Cas12a utilizes a combination of its PAM-interacting (PI) and wedge (WED) domains to recognize the T-rich PAM and initiate DNA unwinding [7] [11]. This structural distinction translates to functional differences: Cas9 creates blunt-ended double-strand breaks approximately 3-4 nucleotides upstream of the PAM, while Cas12a introduces staggered cuts with 5' overhangs distal to the PAM recognition site [2]. The staggered ends produced by Cas12a are particularly advantageous for homology-directed repair (HDR) as they facilitate more predictable repair outcomes and potentially enhance precise gene insertion compared to the blunt ends generated by Cas9.
Table 1: Fundamental Characteristics of Cas9 and Cas12a PAM Recognition
| Feature | CRISPR-Cas9 | CRISPR-Cas12a |
|---|---|---|
| PAM Sequence | 5'-NGG-3' (downstream) | 5'-TTTV-3' (upstream) |
| PAM Length | 3 nucleotides | 4 nucleotides |
| PAM Location | 3' of target sequence | 5' of target sequence |
| Break Type | Blunt ends | Staggered ends (5' overhangs) |
| Guide RNA | crRNA + tracrRNA (or sgRNA) | Single crRNA |
| Natural Multiplexing | Requires engineering | Native capability via crRNA arrays |
Quantifying Targetable Genomic Space
The distinct PAM requirements of Cas9 and Cas12a directly translate to significant differences in their theoretical targeting ranges across various genomes. Quantitative assessments reveal that the canonical SpCas9 with its NGG PAM can access approximately 6% of a typical genome, while wild-type Cas12a with its TTTV PAM is restricted to only about 1% of genomic sites [7]. This substantial disparity stems from the relative abundance of their respective PAM sequences throughout most eukaryotic genomes, with NGG motifs occurring more frequently than TTTV motifs in many organisms.
This limitation in targeting space has prompted extensive engineering efforts to develop Cas12a variants with relaxed PAM requirements. Notably, directed evolution approaches have yielded Flex-Cas12a, a variant carrying six mutations (G146R, R182V, D535G, S551F, D665N, and E795Q) that recognizes 5'-NYHV-3' PAMs (where Y is C or T, H is A, C, or T) [7]. This engineered variant dramatically expands the potential genome accessibility from approximately 1% to over 25%, significantly broadening Cas12a's utility for both therapeutic and agricultural genome engineering applications. Similarly, engineered Cas9 variants with expanded PAM recognition (such as xCas9 and SpCas9-NG) have also been developed, though these were not specifically covered in the search results.
Table 2: Targeting Space and Efficiency Comparison of Cas Nucleases
| Nuclease | PAM Sequence | Theoretical Genomic Coverage | Editing Efficiency | Key Applications |
|---|---|---|---|---|
| SpCas9 | 5'-NGG-3' | ~6% of genome [7] | 90-100% mutagenesis in maize [10] | Broad applications in gene knockout |
| LbCas12a (WT) | 5'-TTTV-3' | ~1% of genome [7] | 0-60% mutagenesis in maize [10] | AT-rich genome targeting |
| Flex-Cas12a | 5'-NYHV-3' | ~25% of genome [7] | Retains efficient cleavage [7] | Expanded locus targeting |
| SaCas9 | 5'-NNGRRT-3' | Smaller than SpCas9 | High efficiency in plants [9] | AAV delivery applications |
Experimental Evidence: Direct Comparative Studies
Maize Genome Editing Efficiency
A direct comparison of Cas9 and Cas12a activities in maize targeting the glossy2 gene revealed significant efficiency differences. Researchers introduced constructs carrying SpCas9-gRNA and LbCas12a-crRNA into maize inbred B104 embryos using Agrobacterium-mediated transformation [10]. On-target mutation analysis demonstrated that 90-100% of the Cas9-edited T0 plants carried indel mutations, with 63-77% being homozygous or biallelic mutants. In stark contrast, only 0-60% of Cas12a-edited T0 plants contained on-target mutations, indicating substantially lower editing efficiency under similar experimental conditions [10].
Further investigation into multiplex gene editing capabilities targeting the Opaque2 (O2) gene in maize demonstrated that the CRISPR/Cpf1 system showed lower editing efficiency than the CRISPR/Cas9 system in the T0 and T1 generations [12]. However, in the T2 generation, the CRISPR/Cpf1 system generated more types of new mutations, suggesting distinct long-term editing dynamics between the two systems. The study also found that editing efficiency in the CRISPR/Cas12a system positively correlated with the expression level of the Cpf1 protein, highlighting the importance of optimization for achieving satisfactory results [12].
Editing Precision and Specificity
Beyond raw efficiency metrics, editing precision represents a critical consideration for research applications. A comparative study in Chlamydomonas reinhardtii examined Cas9 and Cas12a activities by targeting overlapping regions at three independent loci [13]. The research found that while Cas9 and Cas12a ribonucleoproteins (RNPs) co-delivered with ssODN repair templates induced similar levels of total editing (achieving 20-30% in all viably recovered cells), the level of precision editing was slightly higher for Cas12a [13]. This enhanced precision, coupled with Cas12a's inherent capacity for multiplexed editing through its native crRNA processing activity, positions Cas12a as a valuable tool for applications requiring precise genetic modifications.
The specificity of these systems was further evaluated through genome-wide off-target analysis. CIRCLE-seq analysis of potential off-target sites for Cas9 in maize identified 18 and 67 potential off-targets for two different gRNAs, with an average of five mismatches compared to the target sites [10]. Importantly, sequencing analysis of a selected subset of these off-target sites revealed no detectable level of mutations in the T1 plants, suggesting that the CRISPR/Cas9 system used in this study was highly specific for genome editing in maize [10].
Diagram 1: PAM Recognition and Cleavage Mechanisms
Experimental Protocols for PAM Specificity Analysis
Directed Evolution for PAM Relaxation
To address the limited targeting range of wild-type Cas12a, researchers have employed sophisticated directed evolution approaches. One protocol utilized a bacterial-based selection system combined with rational engineering to identify LbCas12a variants with expanded PAM recognition [7]. The methodology involved:
Library Generation: Creating LbCas12a variants with random mutations in the PAM-interacting and wedge domains using error-prone PCR with a controlled error rate (6-9 nucleotide mutations per kilobase) by adding MnCl₂ to the PCR reaction [7].
Dual Selection System: Implementing a chloramphenicol-resistant bacterial expression library with crRNAs designed to direct cleavage at target sequences adjacent to noncanonical PAMs (AGCT, AGTC, TGCA, TCAG) within a ccdB lethal gene located in the selection plasmid [7].
Positive Selection: Electroporating library DNA into E. coli strain BW25141 containing an ampicillin-resistant plasmid encoding an arabinose-inducible ccdB lethal gene, then plating on arabinose and chloramphenicol plates to select for functional PAM-relaxed variants [7].
Variant Characterization: Isolating positive colonies, sequencing plasmids, and performing biochemical and cell-based assays to validate PAM flexibility while retaining recognition of canonical PAMs [7].
This approach yielded Flex-Cas12a, which utilizes 5'-NYHV-3' PAMs and expands DNA recognition sites to approximately 25% of the human genome while maintaining efficient cleavage activity at canonical sites [7].
SELEX-seq for Off-Target Binding Profiling
Understanding the specificity of Cas nucleases requires comprehensive off-target characterization. A modified SELEX-seq (Systematic Evolution of Ligands by Exponential Enrichment with sequencing) approach has been adapted to study Cas12a off-target binding [11]:
Library Design: Constructing a DNA duplex library based on a Cas12a target site containing a TTTG PAM, with a constant target-strand sequence and randomized non-target-strand sequences at 6 protospacer positions adjacent to the PAM [11].
Library Validation: Confirming library functionality with control experiments using catalytically active Cas12a, demonstrating that the DNA library was cleaved with an on-target crRNA guide but not with an off-target guide [11].
Binding Selection: Incubating the library with catalytically-inactive dFnCas12a effector complex, then separating bound and unbound DNA populations using native gel shift assays [11].
Sequencing and Analysis: Recovering and sequencing bound and unbound DNA populations, then computing relative enrichment values to determine sequence preferences for Cas12a binding [11].
This protocol revealed that PAM-adjacent DNA flexibility tunes Cas12a off-target binding, with favorable off-target binding depending on the presence of a properly formed PAM and unpairing of the protospacer at the PAM+1-3 region [11].
Diagram 2: Experimental Workflow for PAM Specificity Studies
The Scientist's Toolkit: Essential Research Reagents
Table 3: Essential Research Reagents for PAM Specificity Studies
| Reagent/Category | Specific Examples | Function/Application | Experimental Considerations |
|---|---|---|---|
| Cas Expression Vectors | pYPQ166 (Cas9 entry vector), pYPQ210 (destination vector) [10] | Stable expression of Cas nucleases in plants | Use strong constitutive promoters (e.g., PZmUbi) for optimal expression |
| Guide RNA Systems | OsU3 promoter for gRNAs, dual-Pol II system for crRNAs [10] | Expression of guide RNAs | Cas12a crRNAs can be processed from arrays for multiplexing |
| Transformation Tools | Agrobacterium strain, bialaphos-resistance (bar) gene [10] [12] | Plant transformation and selection | Efficiency varies by plant genotype and target tissue |
| Editing Analysis Kits | CIRCLE-seq for off-target profiling [10] | Genome-wide off-target identification | More comprehensive than in silico prediction alone |
| Specificity Enhancers | High-fidelity variants (e.g., hfCas12Max) [9] | Reduce off-target effects while maintaining on-target activity | Often involve mutations in REC domains |
| PAM-Relaxed Variants | Flex-Cas12a (G146R, R182V, D535G, S551F, D665N, E795Q) [7] | Expand targetable genomic space | Validate efficiency with new PAMs in your system |
The comparative analysis between Cas9's NGG and Cas12a's TTTV PAM specificities reveals a nuanced landscape for genome editing applications. CRISPR-Cas9, with its broader genomic coverage (~6%) and consistently high editing efficiency across diverse organisms, remains the workhorse for most standard gene knockout applications [10] [7]. However, CRISPR-Cas12a offers distinct advantages for specialized applications, including its native multiplexing capability through crRNA arrays, staggered cuts with 5' overhangs that favor precise HDR, and superior performance in AT-rich genomic regions [2] [12].
For researchers navigating this decision landscape, the experimental evidence suggests a contingency-based approach: Cas9 excels in applications requiring maximum editing efficiency and broad targetability, while Cas12a offers advantages for precise editing, multiplexing, and accessing T-rich genomic regions. The emergence of engineered variants like Flex-Cas12a with dramatically expanded PAM recognition (up to 25% genomic coverage) further blurs the distinction between these systems and promises to unlock new targeting possibilities [7]. As both systems continue to evolve through protein engineering, the optimal choice increasingly depends on specific experimental requirements rather than inherent superiority of one system over the other, empowering researchers with an expanding toolkit for precision genome manipulation.
The ability to simultaneously modify multiple genomic loci, known as multiplex genome editing, has become a cornerstone of advanced genetic research and therapeutic development. This capability is crucial for modeling polygenic diseases, engineering complex metabolic pathways, and understanding gene regulatory networks. The CRISPR-Cas systems, particularly the widely adopted Cas9 and the increasingly prominent Cas12a, have revolutionized this field by providing programmable platforms for precise genetic modifications. While both systems offer distinct advantages, their architectural differences significantly impact their applicability for multiplexed experiments. The fundamental challenge in multiplex editing lies in the efficient co-delivery and processing of multiple guide RNAs to direct nucleases to their respective targets, a process where Cas12a's unique intrinsic ribonuclease (RNase) capability provides a distinct strategic advantage. This article objectively compares the performance of CRISPR-Cas9 and Cas12a for multiplex editing research, with a specific focus on how Cas12a's RNase activity simplifies experimental workflows, enhances efficiency, and enables more sophisticated genetic engineering applications. By examining direct comparative studies and recent experimental data, we provide researchers with a comprehensive evidence-based analysis to inform their genome editing tool selection.
Fundamental Mechanisms: crRNA Biogenesis and Processing
The divergence in guide RNA processing mechanisms between Cas9 and Cas12a represents the foundational difference that dictates their multiplexing capabilities. Understanding these distinct biological pathways is essential for appreciating their respective advantages and limitations in complex genome editing experiments.
Cas9's Dual RNA Requirement: The Class II Type II CRISPR-Cas9 system, derived from Streptococcus pyogenes, requires two RNA components for function: a CRISPR RNA (crRNA) containing the target-specific spacer sequence and a trans-activating crRNA (tracrRNA) that facilitates processing and complex formation [14] [15]. In most laboratory applications, these two elements are combined into a single-guide RNA (sgRNA) through an artificial linkage. However, this engineered solution does not eliminate the fundamental requirement for both functional domains. For multiplex editing with Cas9, each target requires its own complete sgRNA expression cassette, typically driven by individual RNA polymerase III promoters (e.g., U6). This approach creates significant genetic instability due to repetitive promoter sequences and limits packaging capacity in size-constrained delivery vectors like adeno-associated viruses (AAVs) [15].
Cas12a's Autonomous RNase Activity: In contrast, the Class II Type V CRISPR-Cas12a system possesses an inherent endoribonuclease capability that enables autonomous processing of its guide RNAs. Cas12a recognizes and cleaves a long precursor crRNA (pre-crRNA) transcript into mature, functional crRNAs [14]. RNA sequencing analyses from studies in Francisella novicida revealed that mature Cas12a crRNAs are 42-44 nucleotides in length, with the first 19-20 nucleotides forming the direct repeat and the remaining 23-25 nucleotides comprising the spacer sequence [14]. This processing occurs without requiring tracrRNA or additional host factors, as the RNase active site resides within the WED-III subdomain of the Cas12a protein itself [14]. This self-contained processing mechanism forms the basis of Cas12a's advantage in multiplexed applications.
Table 1: Fundamental Comparison of Cas9 and Cas12a Guide RNA Systems
| Feature | Cas9 | Cas12a |
|---|---|---|
| Guide RNA Components | crRNA + tracrRNA (typically fused as sgRNA) | Single crRNA only |
| Guide RNA Length | ~100 nt for sgRNA | ~42-44 nt for mature crRNA |
| Processing Mechanism | Requires host RNase III + tracrRNA | Intrinsic RNase activity in Cas12a protein |
| pre-crRNA Source | Not typically used; individual sgRNAs expressed | Directly expressed and processed |
| Multiplex Guide Delivery | Multiple individual expression cassettes | Single transcript with tandem crRNAs |
Experimental Evidence: Direct Performance Comparison
Recent comparative studies across diverse biological systems provide quantitative evidence of Cas12a's performance in multiplex editing scenarios. These investigations reveal both the advantages and limitations of Cas12a relative to Cas9, offering researchers actionable data for experimental planning.
Editing Efficiency and Patterns: A comprehensive 2024 study in tomato cells directly compared LbCas12a with SpCas9 by investigating on-target efficacy and specificity at 35 overlapping target sites [4]. The research found that LbCas12a was an efficient, robust addition to SpCas9, with similar overall editing efficiencies, though these were target-dependent. Notably, the mutational patterns differed significantly—LbCas12a induced more and larger deletions than SpCas9, which can be advantageous for specific genome editing applications where complete gene knockout is desired [4]. This characteristic may be attributed to the staggered DNA ends with 5' overhangs generated by Cas12a, compared to the blunt ends produced by Cas9, potentially leading to different repair outcomes through non-homologous end joining or microhomology-mediated end joining pathways.
Precision in Genome Editing: When comparing Cas9 and Cas12a for gene editing in Chlamydomonas reinhardtii, researchers found that while both systems achieved 20-30% total editing in all viably recovered cells when delivered as ribonucleoproteins with ssODN repair templates, Cas12a demonstrated slightly higher precision in ssODN-templated genome editing [13]. However, the same study noted that Cas9 targeted more sites and induced more total edits when RNPs were delivered alone, without repair templates, making Cas9 the preferable enzyme for certain genome engineering applications in this model organism [13]. This highlights the importance of matching the nuclease characteristics to specific experimental goals.
Multiplexing Performance in Fungal Systems: The implementation of Cas12a in Aspergillus niger demonstrated its robust capability for complex multiplexed editing. Researchers developed a toolbox that enabled assembly of plasmids containing gRNA multiplex expression cassettes capable of expressing a multi-gRNA precursor processed via Cas12a's RNase activity [16]. Using this system, they successfully constructed vectors delivering up to ten gRNAs and demonstrated that three simultaneous deletions could be introduced robustly by targeting each gene with several gRNAs, without prior gRNA validation or genomically integrated selection markers [16]. This streamlined approach allowed multiple specific genomic modifications to be performed in less than two weeks, significantly accelerating strain construction efforts.
Table 2: Experimental Performance Comparison of Cas9 and Cas12a in Multiplex Editing
| Parameter | Cas9 | Cas12a | Experimental Context |
|---|---|---|---|
| Overall Editing Efficiency | Target-dependent; generally high | Similar overall efficiency; target-dependent | Tomato protoplasts [4] |
| Editing Precision | High | Slightly higher precision with ssODN templates | Chlamydomonas reinhardtii [13] |
| Mutation Pattern | Blunt ends; typical indels | Staggered ends; more & larger deletions | Tomato cells [4] |
| Multiplex Capacity | Limited by delivery of multiple sgRNAs | Up to 10 gRNAs demonstrated | Aspergillus niger [16] |
| Off-target Activity | Well-characterized off-target concerns | Fewer off-targets; mismatches tolerated in distal region | Tomato; off-targets with 1-2 mismatches distal from PAM [4] |
Cas12a Engineering Advances: Enhanced Efficiency and Specificity
The initial adoption of Cas12a in mammalian systems was hampered by variable editing efficiencies compared to Cas9. However, recent protein engineering efforts have yielded dramatically improved Cas12a variants that retain its native advantages while overcoming early limitations.
Hyper-Efficient Cas12a Variants: Through structure-guided protein engineering, researchers have developed hyper-efficient LbCas12a variants, termed hyperCas12a, with significantly enhanced efficacy [8]. The engineering approach focused on mutating negatively charged residues within 10Å of the target DNA to positively charged arginines, potentially increasing binding affinity to negatively-charged DNA [8]. A quadruple mutant (D156R/D235R/E292R/D350R) demonstrated remarkable performance improvements, particularly under low crRNA conditions relevant to in vivo applications where molecular concentrations are limiting [8]. This hyperCas12a variant achieved ~60-fold higher activation than wildtype Cas12a when using RNA polymerase II-driven crRNA expression, and outperformed wildtype by >300-fold under limiting crRNA conditions [8].
High-Fidelity Variants: Engineering efforts have also addressed specificity concerns through high-fidelity Cas12a variants. The enhanced AsCas12a-HF1 (enAsCas12a-HF1) with substitutions E174R/N282A/S542R/K548R has demonstrated improved targeting accuracy while maintaining efficiency [17]. These engineered variants have been successfully incorporated into knock-in mouse models, enabling efficient multiplexed genome engineering in vivo without discernible pathology [17]. The development of these advanced Cas12a variants has effectively addressed earlier efficiency concerns while preserving the innate multiplexing advantages of the system.
Practical Implementation: Methodologies and Reagent Systems
Translating the theoretical advantages of Cas12a into practical experimental success requires appropriate methodological approaches and reagent systems. Below, we detail key protocols and resources that leverage Cas12a's unique capabilities for multiplex genome editing.
Detailed Method: Multiplexed Gene Editing in Aspergillus niger Using Cas12a
This protocol demonstrates a robust system for simultaneous modification of multiple genomic loci in fungi, leveraging Cas12a's intrinsic RNase activity for crRNA processing [16].
Experimental Workflow:
- gRNA Array Design: Design 36-44 nt crRNAs targeting each locus of interest, ensuring each contains the 19-20 nt direct repeat followed by the target-specific spacer. Maintain the TTTV PAM requirement in the genomic target sequence.
Vector Assembly: Assemble the multiplex gRNA expression cassette using Golden Gate or USER cloning approaches. The system employs bio-blocks with complementary overhangs to sequentially assemble up to 10 gRNAs into a single expression cassette flanked by tRNA sequences for initial processing by cellular RNases.
Transformation: Deliver the assembled CRISPR plasmid containing the Cas12a endonuclease and gRNA array expression cassette to A. niger protoplasts using standard fungal transformation techniques. Selection is performed using appropriate nutritional or antibiotic markers.
Screening and Validation: Screen transformants for successful gene editing events using a combination of PCR screening and sequencing. The efficiency of multiplex editing is quantified by calculating the percentage of transformants with modifications at all targeted loci.
Key Advantages of This System:
- Eliminates the need for individual gRNA validation through the use of multiple gRNAs per target
- Enables complex genetic engineering projects with timeframes under two weeks
- Allows marker-free modifications through direct screening of editing events
- Demonstrates robust simultaneous deletion of three distinct genomic regions
The Scientist's Toolkit: Essential Reagents for Cas12a Multiplexing
Table 3: Key Research Reagent Solutions for Cas12a Multiplex Editing
| Reagent / Resource | Function / Application | Example Source / System |
|---|---|---|
| LbCas12a Expression Vector | Codon-optimized Cas12a with nuclear localization signals | Addgene #69976 [4] |
| enAsCas12a-HF1 | High-fidelity engineered Cas12a variant with enhanced specificity | Knock-in mouse models [17] |
| hyperCas12a Variant | Engineered LbCas12a with enhanced activity under low crRNA conditions | Structure-guided mutants (D156R/D235R/E292R/D350R) [8] |
| Golden Gate Cloning System | Modular assembly of crRNA arrays with standardized parts | MoClo toolkit & Plant Parts kit [4] |
| Cas12a-knock-in Mice | In vivo model with constitutive Cas12a expression for multiplex editing | Rosa26 locus knock-in [17] |
| crRNA Array Expression Cassette | Single transcript for multiple gRNAs with direct repeats | Polymerase II promoter systems [8] |
Application Scope: From Basic Research to Therapeutic Development
The unique capabilities of Cas12a in multiplexed editing have enabled applications across diverse biological systems and research domains, demonstrating its versatility and practical utility.
In Vivo Disease Modeling: The development of Cas12a-knock-in mice has facilitated sophisticated in vivo disease modeling through multiplexed genome engineering. Researchers have demonstrated efficient quadruplex gene knockout in vivo using a single AAV vector simultaneously targeting murine Trp53, Apc, Pten, and Rb1, resulting in rapid induction of salivary gland squamous cell carcinoma and lung adenocarcinoma [17]. This approach enables more accurate modeling of complex cancer genotypes that require simultaneous perturbation of multiple genes, better recapitulating the polygenic nature of human malignancies.
Combinatorial Genetic Screening: Cas12a's multiplexing capabilities make it ideally suited for combinatorial genetic screening applications. The ability to deliver numerous gRNAs from a single transcript enables more efficient exploration of genetic interactions, including epistasis, redundancy, synergy, and antagonism [17]. This is particularly valuable for deciphering complex gene regulatory networks and identifying synthetic lethal interactions for therapeutic development.
Cell Engineering and Gene Therapy: In the realm of cell engineering, Cas12a has shown significant promise for immune cell engineering. The system has been utilized for retrovirus-based engineering of CD4+ and CD8+ T cells, B cells, and bone-marrow-derived dendritic cells, enabling both knock-in of chimeric antigen receptors and simultaneous knockout of checkpoint inhibitors [17]. This simultaneous multiplex editing capability streamlines the production of engineered therapeutic cells for advanced immunotherapies.
The experimental evidence comprehensively demonstrates that Cas12a's intrinsic RNase activity and simplified guide RNA architecture provide distinct advantages for multiplex genome editing applications. The ability to process multiple crRNAs from a single transcript significantly streamlines vector construction, enhances delivery efficiency, and enables more complex genetic perturbations than possible with Cas9-based systems. While Cas9 remains a powerful tool for many applications, particularly those requiring maximum on-target efficiency at individual loci, Cas12a offers superior capabilities for experiments requiring simultaneous modification of multiple genomic targets. Recent protein engineering efforts have addressed early efficiency concerns, with hyper-efficient and high-fidelity variants now delivering robust performance across diverse biological systems. Researchers should consider Cas12a as the preferred platform for sophisticated multiplex editing applications including complex disease modeling, combinatorial genetic screening, and therapeutic cell engineering, where its crRNA processing advantage translates to tangible experimental benefits.
The CRISPR-Cas revolution in genome engineering has been largely powered by the Cas9 nuclease. However, the discovery and adaptation of Cas12a have provided scientists with a distinct and powerful alternative. The fundamental difference in the DNA break profiles generated by these two nucleases—blunt ends from Cas9 versus staggered ends from Cas12a—has profound implications for experimental design and outcomes in multiplex editing research [2]. This guide provides an objective, data-driven comparison of these systems, focusing on their mechanisms, editing efficiencies, mutational patterns, and practical applications to inform tool selection for specific research goals.
Molecular Mechanisms and DNA Break Anatomy
The initial step of CRISPR-Cas targeting is defined by the Protospacer Adjacent Motif (PAM), a short DNA sequence adjacent to the target site that the nuclease must recognize to initiate binding.
- Cas9 PAM (Type II System): The commonly used Streptococcus pyogenes Cas9 requires a 5'-NGG-3' PAM located upstream of the target site on the non-target DNA strand [18] [10]. This G-rich PAM can sometimes restrict targeting in AT-rich genomic regions.
- Cas12a PAM (Type V System): Cas12a recognizes a 5'-TTTV-3' PAM (where V is A, C, or G) located downstream of the target site [18] [2]. This T-rich PAM expands the targeting space, particularly in promoter regions and other AT-rich sequences that are challenging for Cas9.
Once bound, the nucleases catalyze double-strand breaks (DSBs), but the nature of these breaks is fundamentally different, as shown in the following diagram.
- Cas9 creates blunt ends: The Cas9 nuclease makes a clean cut through both DNA strands at the same position, typically 3 base pairs upstream of the PAM site, resulting in double-stranded DNA fragments with no unpaired nucleotides [18] [2].
- Cas12a creates staggered ends: Cas12a induces DSBs with 5' overhangs of 4-5 nucleotides. The cuts are typically staggered, with one strand being cut 18 nucleotides downstream of the PAM and the other 23 nucleotides downstream, creating "sticky ends" [2] [4].
Quantitative Comparison of Editing Outcomes
The structural differences in DNA breaks lead to distinct repair profiles by the cell's endogenous DNA repair machinery, primarily the Non-Homologous End Joining (NHEJ) pathway. The table below summarizes the key comparative characteristics of the two systems.
Table 1: Fundamental Characteristics of Cas9 and Cas12a
| Feature | CRISPR-Cas9 | CRISPR-Cas12a |
|---|---|---|
| Nuclease Class | Type II | Type V |
| PAM Sequence | 5'-NGG-3' (G-rich) [18] | 5'-TTTV-3' (T-rich) [18] |
| PAM Location | 3' upstream of target [18] | 5' downstream of target [18] |
| DNA Break Type | Blunt ends [2] | Staggered ends with 5' overhangs [2] [4] |
| Guide RNA | Two-part system (crRNA + tracrRNA) or single chimeric sgRNA [2] [10] | Single, shorter crRNA (~42-44 nt) [2] |
| Multiplexing | Requires multiple or arrayed gRNAs | Native processing of crRNA arrays due to RNase activity [17] |
These mechanistic differences translate into measurable variations in editing outcomes. Research across various plant and algal models provides quantitative data on their performance.
Table 2: Comparative Editing Performance of Cas9 and Cas12a
| Organism / Experiment | Editing Efficiency | Mutational Pattern (Indel Profile) | Key Findings |
|---|---|---|---|
| Rice (RNP Delivery) [18] | LbCas12a > WT Cas9 | Cas9: 1-2 bp indels or 20-30 bp deletionsLbCas12a: 2-20 bp deletions without PAM loss | LbCas12a RNP achieved higher mutagenesis frequency at the single OsPDS target site evaluated. |
| Tomato Protoplasts [4] | Similar overall, but target-dependent | Cas9: Primarily small indelsLbCas12a: Significantly more and larger deletions | Cas12a's tendency for larger deletions can be advantageous for gene knock-outs. |
| Chlamydomonas reinhardtii [13] | Cas9 RNPs alone induced more total edits than Cas12a RNPs | Cas12a: Showed slightly higher precision when using ssODN repair templates | With ssODN templates, total editing levels were similar, but Cas12a was more precise. |
| Maize [10] | Cas9: 90-100% of T0 plants had mutationsCas12a: 0-60% of T0 plants had mutations | Not detailed | The specific Cas9 system used was more efficient than the Cas12a system in this study. |
Detailed Experimental Protocols and Data
To ensure the reproducibility of comparative studies, this section outlines the core methodologies used to generate the data cited in this guide.
Ribonucleoprotein (RNP) Delivery in Rice
A direct comparison of Cas9 and Cas12a was performed in rice using RNP complexes to ensure transient editing activity and avoid DNA integration [18].
- Protein and Guide RNA: Purified WT Cas9, HiFi Cas9, and LbCas12a proteins were complexed with chemically synthesized guide RNAs (crRNA1 for Cas9, crRNA3 for Cas12a) targeting the same sequence in the OsPDS gene [18].
- Delivery and Selection: RNP complexes were co-delivered with a plasmid (pCAMBIA1301) into 5-day-old mature seed-derived rice embryos via biolistic transformation. Transformed cells were selected and analyzed [18].
- Genotyping and Phenotyping: Mutation efficiency was determined by tracking the albino phenotype caused by bi-allelic PDS knockout and confirmed by molecular genotyping to characterize the nature of the induced indels [18].
High-Throughput Amplicon Sequencing in Tomato Protoplasts
A highly quantitative comparison was conducted in tomato protoplasts, allowing for the screening of numerous gRNAs/crRNAs with deep sequencing [4].
- Vector Construction: A Golden Gate-based system was used to clone expression vectors for human-codon-optimized LbCas12a and SpCas9, along with their respective guide RNAs [4].
- Target Site Selection: 35 overlapping target sites within the bHLH transcription factor gene family were selected to allow a direct comparison of both nucleases at the same genomic locations [4].
- Transfection and Analysis: Tomato protoplasts were transfected with the CRISPR constructs. After 48 hours, DNA was extracted, on-target and predicted off-target sites were amplified, and the products were subjected to high-throughput amplicon sequencing to quantify editing efficiency and profile the resulting mutations [4].
The workflow for such a comparative analysis is standardized as follows.
The Scientist's Toolkit: Essential Reagents
Successful genome editing requires carefully selected molecular tools. The table below lists key reagents used in the featured experiments.
Table 3: Key Research Reagent Solutions for CRISPR-Cas Comparative Studies
| Reagent / Material | Function in Experiment | Example from Cited Studies |
|---|---|---|
| Purified Cas Protein (RNP) | Direct delivery of editing machinery; transient activity with reduced off-targets and cytotoxicity [18] [19]. | LbCas12a, AsCas12a, WT SpCas9, HiFi SpCas9 proteins [18] [19]. |
| Chemically Synthesized Guide RNA | High-purity, consistent guide RNA for RNP complex formation or for assessing in vitro activity. | 36-41 nt crRNAs for Cas9 and Cas12a targeting rice OsPDS [18]. |
| Golden Gate MoClo Toolkit | Modular, efficient cloning system for assembling expression vectors for nucleases and guide RNA arrays [4]. | Used to construct Cas12a nuclease and crRNA expression vectors for tomato protoplast assays [4]. |
| Codon-Optimized Nuclease Genes | Enhanced expression of the Cas nuclease in the host organism of interest, improving editing efficiency. | Human-codon-optimized LbCas12a, AsCas12a, FnCas12a for expression in tomato protoplasts [4]. |
| Plasmid Donor DNA | Provides a template for Homology-Directed Repair (HDR) or for selecting transformed cells. | Plasmid pCAMBIA1301 used as a co-delivery selectable marker in rice RNP experiments [18]. |
Discussion and Research Implications
The choice between Cas9 and Cas12a is not a matter of superiority but of strategic selection based on the specific research objective.
- Selecting Cas9 is advantageous when: The target site is flanked by an NGG PAM, the goal is to induce a high frequency of small indels for gene disruption, or when a proven, highly efficient system with extensive published validation is required.
- Selecting Cas12a is advantageous when: Targeting AT-rich genomic regions [2], performing multiplexed genome editing using a single crRNA array [17], aiming for larger deletions for complete gene knock-outs [4], or when the staggered ends are desired to facilitate specific HDR or MMEJ repair outcomes [4].
For researchers working with advanced in vivo models, the recent development of Cas12a-knock-in mice provides a powerful toolkit for multiplexed genome editing, disease modeling, and immune-cell engineering without the delivery constraints associated with the large size of Cas12a vectors [17].
In conclusion, the blunt ends of Cas9 and the staggered cuts of Cas12a offer complementary paths for genome engineering. A deep understanding of their contrasting DNA break profiles empowers researchers to make informed decisions, optimizing the success and efficiency of their multiplex editing projects.
Implementing Multiplex Editing: From Immune Cell Engineering to In Vivo Modeling
Designing Single-Transcript crRNA Arrays for Efficient Multi-Gene Knockout
The ability to simultaneously modify multiple genomic loci—multiplex gene editing—has become a cornerstone of modern genetic research, enabling the study of complex gene networks, synthetic lethal interactions, and polygenic diseases. The CRISPR-Cas system has emerged as the most powerful platform for multiplex editing due to its simplicity and programmability. While CRISPR-Cas9 has been widely adopted since its discovery, the more recently characterized CRISPR-Cas12a system offers distinct advantages for multiplexing applications due to its unique enzymatic properties and simpler array architecture [14]. This guide provides an objective comparison between these two systems, focusing specifically on their application in designing single-transcript crRNA arrays for efficient multi-gene knockout experiments.
The fundamental difference in their approach to multiplexing stems from their crRNA biogenesis mechanisms. Cas9 requires either multiple separate guide RNA expression cassettes or additional processing elements such as ribozymes or tRNA sequences, while Cas12a possesses inherent RNase activity that enables it to process a single long transcript into multiple mature crRNAs [14]. This intrinsic capability makes Cas12a particularly suitable for constructing compact, efficient single-transcript crRNA arrays, simplifying vector design and potentially increasing editing efficiency in complex multi-gene knockout experiments.
Mechanistic Comparison: Cas9 vs. Cas12a for Multiplex Editing
Fundamental System Architectures
The CRISPR-Cas9 and CRISPR-Cas12a systems differ significantly in their molecular architectures and mechanisms, which directly impacts their application in multiplex gene editing. The CRISPR-Cas9 system utilizes a dual RNA structure consisting of a CRISPR RNA (crRNA) for target recognition and a trans-activating crRNA (tracrRNA) that is essential for processing and maturation, though these are typically combined into a single-guide RNA (sgRNA) for experimental applications [10]. For multiplex editing with Cas9, researchers must express multiple sgRNAs, which often requires complex vector designs with repeated promoter sequences or additional processing elements such as ribozymes or tRNA sequences to liberate individual guides from a single transcript [12].
In contrast, the CRISPR-Cas12a system requires only a single CRISPR RNA (crRNA) for both processing and targeting functions. Cas12a possesses intrinsic RNase activity that enables it to process its own pre-crRNA into mature crRNAs without requiring additional cellular factors [14]. This key enzymatic difference allows Cas12a to natively process a single transcript containing multiple crRNAs separated by direct repeats, making it inherently suitable for multiplexed genome editing applications. The Cas12a system recognizes T-rich PAM sequences (5'-TTTV-3', where V is A, C, or G), which expands the targeting scope compared to the G-rich PAM (5'-NGG-3') required by the most commonly used Streptococcus pyogenes Cas9 [10] [14]. Furthermore, Cas12a creates staggered DNA double-strand breaks with 4-5 nucleotide 5' overhangs, unlike the blunt ends generated by Cas9, potentially facilitating specific editing outcomes in multi-gene knockout experiments [14].
crRNA Array Processing Mechanisms
The differential RNA processing mechanisms between Cas9 and Cas12a significantly impact the design of single-transcript arrays for multi-gene knockout experiments. Cas9 lacks intrinsic processing capability for multiple guides from a single transcript, requiring the incorporation of exogenous processing elements such as tRNA or ribozyme sequences between individual sgRNAs. These additional elements increase the size and complexity of the multiplex construct and may introduce variability in processing efficiency between different guide positions within the array [12].
In contrast, Cas12a autonomously processes pre-crRNA through its RNase activity, recognizing and cleaving within the direct repeat sequences that separate individual crRNA units in a polycistronic transcript [16] [14]. This self-contained processing mechanism enables more compact array designs and theoretically more consistent maturation of individual crRNAs. The simplicity of the Cas12a array architecture—consisting essentially of alternating direct repeats and spacer sequences—facilitates the construction of complex multi-guide expression cassettes, as demonstrated in Aspergillus niger where researchers successfully assembled arrays containing up to ten crRNAs using a modular bio-block system [16].
Performance Comparison: Experimental Data Across Biological Systems
Editing Efficiency in Plant Systems
Comparative studies in maize have provided valuable quantitative data on the performance of Cas9 versus Cas12a for multiplex genome editing. When targeting the glossy2 (gl2) gene with overlapping target sites, Cas9 demonstrated superior editing efficiency in initial generations, with 90-100% of T0 plants carrying indel mutations and 63-77% being homozygous or biallelic mutants [10]. In contrast, Cas12a showed considerably lower efficiency in the same experimental system, with only 0-60% of T0 plants exhibiting on-target mutations [10].
However, this efficiency gap appears to narrow in subsequent generations. A separate maize study targeting the Opaque2 (O2) gene found that while Cas9 maintained higher efficiency in T0 and T1 generations, Cas12a generated more diverse mutation types in T2 plants, with a tendency to create edits between target sites rather than within the immediate on-target region [12]. This suggests potential trade-offs between initial editing efficiency and the diversity of editing outcomes, which may be relevant for different research applications.
Table 1: Performance Comparison of Cas9 and Cas12a in Maize Multiplex Editing
| Parameter | CRISPR-Cas9 | CRISPR-Cas12a | Experimental Context |
|---|---|---|---|
| Editing Efficiency (T0) | 90-100% of plants with indels [10] | 0-60% of plants with indels [10] | gl2 gene targeting in maize |
| Homozygous/Biallelic (T0) | 63-77% [10] | Not reported | gl2 gene targeting in maize |
| Mutation Diversity (T2) | Limited, on-target editing preferred [12] | Higher, editing between targets [12] | O2 gene targeting in maize |
| Expression Correlation | Not strongly correlated with efficiency | Positive correlation with Cpf1 expression [12] | O2 gene targeting in maize |
Efficiency in Fungal and Mammalian Systems
Beyond plant systems, Cas12a has demonstrated robust performance in various biological contexts. In Aspergillus niger, researchers developed a sophisticated CRISPR-Cpf1 toolbox that enabled simultaneous targeting of three different genes using nine gRNAs (three per gene) delivered as a single precursor RNA molecule [16]. This approach achieved robust editing without prior gRNA validation, highlighting the utility of Cas12a's multi-guide array system for complex genome engineering applications in fungi.
In mammalian systems, the development of Cas12a-knock-in mice has facilitated efficient multiplexed genome engineering across diverse cell types and tissues [17]. These models have enabled retrovirus-based immune cell engineering, autochthonous cancer modeling through delivery of multiple CRISPR RNAs as a single array via adeno-associated viruses, and targeted genome editing in liver tissue using lipid nanoparticles. The constitutive expression of Cas12a in these mice did not lead to discernible pathology, supporting the biological safety of Cas12a-based approaches [17].
Table 2: Cas12a Performance Across Different Biological Systems
| Organism/System | Editing Efficiency | Multiplex Capacity | Key Findings |
|---|---|---|---|
| Aspergillus niger [16] | Robust simultaneous deletion of 3 genes | 9 gRNAs (3 per gene) | No prior gRNA validation needed; time frame <2 weeks |
| Mouse primary immune cells [17] | Efficient gene editing across multiple cell types | Quadruplex knockout (4 genes) | Enabled cancer modeling with single AAV vector |
| Kluyveromyces marxianus [20] | 50-100% single-gene knockout; 85-95% knock-in | Triple-fragment knock-in | Chromosomal integration enabled large fragment integration |
| Human cells [21] | Comparable or improved vs. SpCas9 | Not specified | AI-designed editors (OpenCRISPR-1) showed enhanced properties |
Experimental Protocols for Array Design and Validation
Designing and Constructing Cas12a crRNA Arrays
The construction of functional crRNA arrays for Cas12a-mediated multi-gene knockout follows a systematic workflow that leverages the innate processing capabilities of the Cas12a system. The first critical step involves bioinformatic gRNA selection with attention to the TTTV PAM requirement and minimization of potential off-target effects through genome-wide specificity analysis [10]. Tools such as CINDEL and CRISPR-DT can provide predictions of targeting efficacy for individual crRNAs, though their predictive value should be empirically validated [10].
For array assembly, the modular bio-block approach has proven effective, as demonstrated in fungal systems [16]. This method involves designing dsDNA oligonucleotides (bio-blocks) containing individual crRNA units with complementary single-stranded overhangs that facilitate sequential fusion into a complete multi-gRNA expression cassette. The crRNA units are separated by the 19-20 nt direct repeat sequences that Cas12a recognizes and cleaves during processing. The final array is typically cloned between a Pol III promoter (such as U6) and terminator sequence, though some systems have successfully used Pol II promoters for expression of both Cas12a and crRNAs [10] [16].
A key consideration in array design is position-dependent efficiency variation, which may affect editing outcomes at different target loci. While Cas12a's processive crRNA maturation potentially reduces position effects compared to tRNA-or ribozyme-based Cas9 systems, empirical validation of array functionality remains essential. For critical applications where complete knockout of all targets is required, incorporating multiple crRNAs per target gene—as demonstrated in the Aspergillus niger study with three gRNAs per gene—can significantly increase the probability of successful editing at each locus [16].
Delivery and Validation Methods
The delivery method for Cas12a crRNA arrays significantly influences editing outcomes and must be matched to the experimental system. For mammalian cells, viral delivery systems such as adeno-associated viruses (AAVs) have successfully delivered crRNA arrays for in vivo cancer modeling, with one study demonstrating quadruplex knockout of Trp53, Apc, Pten, and Rb1 using a single AAV vector [17]. Lipid nanoparticles (LNPs) have emerged as another promising delivery modality, particularly for therapeutic applications, as they avoid the immunogenicity concerns associated with viral vectors and enable potential redosing [22] [17].
Following delivery, comprehensive molecular validation of editing efficiency and specificity is essential. This typically involves PCR amplification of target regions followed by sequencing to quantify indel frequencies and characterize mutation patterns. For multi-gene knockout experiments, tracking of deletion patterns between target sites is particularly important, as Cas12a has demonstrated a tendency to create edits between—not just within—target sequences in extended generations [12]. Additionally, assessment of protein-level knockout through Western blot or flow cytometry provides functional validation of successful gene disruption, especially in cases where in-frame mutations might preserve partial function.
Advanced Applications and Future Directions
Innovative Applications of Cas12a Multiplex Editing
The unique properties of Cas12a crRNA arrays have enabled sophisticated applications beyond standard gene knockouts. The DAKO (Dual-gene Activation and Knockout) system combines Cas12a-mediated knockout with simultaneous gene activation by integrating Cas12a-knock-in mice with CRISPR activation transgenic lines, enabling complex genetic interaction studies [17]. This approach demonstrates how Cas12a's multiplexing capability can be integrated with other CRISPR technologies for sophisticated perturbation studies.
In metabolic engineering, Cas12a arrays have facilitated combinatorial pathway optimization in industrial microorganisms. In Kluyveromyces marxianus, researchers used a CRISPR-Cas12a system to sequentially knock out multiple succinate dehydrogenase genes, redirecting carbon flux toward succinic acid production and creating a thermotolerant production strain capable of generating the highest reported titer in this species [20]. The efficiency of this system—achieving 50-100% knockout efficiency even with short homology arms—highlights the practical utility of Cas12a for complex strain engineering projects.
Emerging Enhancements and AI-Driven Design
Recent advances in Cas12a engineering have addressed initial limitations and expanded the system's capabilities. The development of enhanced Cas12a variants such as enAsCas12a-HF1, which incorporates E174R/N282A/S542R/K548R substitutions, has improved both editing efficiency and specificity while expanding the PAM recognition repertoire [17]. These engineered variants maintain the multiplexing advantages of wild-type Cas12a while offering improved performance characteristics.
Perhaps the most promising development comes from AI-driven protein design approaches. Researchers have now used large language models trained on diverse CRISPR-Cas sequences to generate novel Cas proteins with optimal properties, including the OpenCRISPR-1 editor that exhibits comparable or improved activity and specificity relative to SpCas9 despite being 400 mutations away in sequence [21]. This AI-driven approach to editor design may yield future Cas12a variants with further enhanced suitability for multiplex gene editing applications.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Cas12a crRNA Array Experiments
| Reagent Category | Specific Examples | Function & Application Notes |
|---|---|---|
| Cas12a Variants | LbCas12a, AsCas12a, enAsCas12a-HF1 [17] | Engineered versions offer improved specificity and expanded PAM recognition |
| Delivery Systems | AAV vectors, Lipid Nanoparticles (LNPs), Retroviral vectors [22] [17] | LNPs enable redosing and avoid viral vector immunogenicity concerns |
| Assembly Systems | Modular bio-blocks, USER cloning, Golden Gate assembly [16] | Modular approaches facilitate construction of complex multi-guide arrays |
| Promoter Systems | U6/U3 Pol III promoters, CAG promoter (mice) [16] [17] | Constitutive or inducible expression depending on experimental needs |
| Validation Tools | CIRCLE-seq, NGS analysis, Western blot, flow cytometry [10] [17] | Multi-level validation from DNA sequencing to protein-level assessment |
| Cell/Animal Models | Cas12a-knock-in mice [17], Δku70 strains (fungi) [16] | Engineered hosts expressing Cas12a simplify delivery requirements |
The strategic implementation of single-transcript crRNA arrays for multi-gene knockout experiments requires careful consideration of both system-specific capabilities and experimental requirements. The CRISPR-Cas12a system offers distinct advantages for multiplexing applications due to its simplified array architecture, inherent processing capability, and expanding toolkit of engineered variants. While CRISPR-Cas9 may currently offer higher editing efficiency in some contexts, particularly in plant systems, Cas12a's unique features—including staggered cut ends, T-rich PAM recognition, and compact array design—make it increasingly valuable for complex genome engineering projects.
The choice between these systems ultimately depends on specific experimental priorities: Cas9 for maximum initial editing efficiency or when G-rich PAM sequences are preferred, versus Cas12a for streamlined multiplex array construction, diverse mutation profiles, or when T-rich PAM sequences are more suitable. As both systems continue to evolve through protein engineering and AI-assisted design, researchers can expect further blurring of current performance boundaries while gaining access to increasingly specialized tools for multi-gene knockout applications.
The advent of clustered regularly interspaced short palindromic repeats (CRISPR) technology has revolutionized genetic engineering, with CRISPR-Cas9 serving as the foundational tool for genome editing in numerous applications. However, for the specialized field of ex vivo immune cell engineering, particularly for developing next-generation cell therapies, CRISPR-Cas12a (Cpf1) has emerged as a powerful alternative with distinct advantages for complex editing tasks. Cas12a offers unique capabilities for multiplexed genome editing, a critical requirement for sophisticated engineering of primary human T-cells and other immune cells to enhance cancer immunotherapy and treat other diseases. This guide objectively compares the performance of Cas12a against Cas9, supported by recent experimental data, providing researchers and drug development professionals with a clear framework for selecting the appropriate CRISPR system for their ex vivo cell engineering projects.
Fundamental System Differences and Advantages for Multiplexing
Cas12a and Cas9, both classified as Class 2 CRISPR systems, originate from different bacterial ancestors and have evolved distinct molecular mechanisms. These fundamental differences directly impact their practical application in multiplexed ex vivo engineering.
- Guide RNA Architecture: Unlike Cas9, which requires a two-part guide system (crRNA and tracrRNA, often fused into a single-guide RNA), Cas12a operates with a single, shorter crRNA (approximately 41-44 nt) and does not require a tracrRNA [2] [12]. This simpler architecture reduces the genetic payload, which is advantageous when packaging into delivery vectors with limited capacity.
- Cleavage Mechanism: Cas9 generates blunt-ended double-strand breaks (DSBs), while Cas12a creates staggered cuts with 5′ overhangs [2] [4]. These "sticky ends" can potentially enhance the efficiency of certain gene insertion strategies by facilitating microhomology-mediated repair pathways.
- Protospacer Adjacent Motif (PAM) Recognition: A key differentiator is the PAM sequence required for target recognition. Cas9 predominantly recognizes G-rich PAM sequences (5'-NGG-3'), whereas Cas12a recognizes T-rich PAMs (5'-TTTV-3', where V is A, C, or G) [23] [2]. This makes Cas12a exceptionally suited for targeting AT-rich genomic regions that are often inaccessible to Cas9, thereby significantly expanding the targetable genome space [2].
- Inherent Multiplexing Capability: Cas12a possesses RNase activity that allows it to process a single long transcript into multiple mature crRNAs [17] [24] [12]. This innate ability to handle crRNA arrays simplifies the design and delivery of systems targeting multiple genes simultaneously, a common requirement in complex immune cell engineering.
Table 1: Fundamental Comparison of Cas12a and Cas9 Systems
| Feature | Cas12a (Cpf1) | Cas9 |
|---|---|---|
| Guide RNA | Single crRNA (~41-44 nt) | sgRNA (crRNA + tracrRNA, ~100 nt) |
| RNA Requirement | tracrRNA-independent | Requires tracrRNA |
| Cleavage Type | Staggered cut with 5' overhang | Blunt-ended cut |
| PAM Sequence | 5'-TTTV-3' (T-rich) | 5'-NGG-3' (G-rich) |
| Multiplexing | Natural crRNA processing from a single array | Requires engineered systems (tRNA, ribozymes, etc.) |
| Key Advantage for Immune Cells | Simplified delivery of multi-gene edits | Widespread validation and high efficiency for single edits |
Diagram 1: CRISPR System Selection for Immune Cell Engineering
Performance Data in Ex Vivo Immune Cell Engineering
Direct comparative studies and platform-specific developments highlight the performance characteristics of Cas12a in primary immune cells, which are often recalcitrant to efficient editing.
Editing Efficiency and Multiplexed Knock-in
A groundbreaking application of Cas12a for T-cell engineering is the CLASH (CRISPR-based Library-scale AAV perturbation with simultaneous HDR knock-in) platform. This system uses Cas12a mRNA electroporation combined with pooled adeno-associated viruses (AAVs) to enable massively parallel knock-in of chimeric antigen receptor (CAR) transgenes into primary human T-cells. In proof-of-concept experiments, CLASH achieved on-target integration efficiencies of 37.4% to 51% in primary human CD8+ T-cells, successfully generating a diverse pool of CAR-T variants for unbiased functional selection [25]. This demonstrates Cas12a's capability for large-scale, complex editing workflows that are essential for advanced cell therapy development.
Further evidence from the development of Cas12a-knock-in mouse models confirms the enzyme's utility in primary immune cells. Researchers reported efficient multiplexed genome engineering in ex vivo derived primary CD4+ and CD8+ T cells, B cells, and bone-marrow-derived dendritic cells (BMDCs) from these mice, observing both DNA-level editing and protein-level reduction of target genes [17] [26]. The constitutive expression of Cas12a in these models did not lead to discernible pathology, supporting its safety for ex vivo applications [17].
Comparative Editing Efficiencies: Cas12a vs. Cas9
While direct head-to-head comparisons in primary human T-cells are limited, data from other biological systems provide insights into their relative performances. In a study in Aspergillus niger, Cas12a demonstrated potential superiority in editing efficiency when a single guide RNA was used, achieving 86.5% efficiency compared to 31.7% for Cas9 at the same locus [23]. However, when two gRNAs were employed for targeting, both systems achieved up to 100% editing efficiency [23]. This suggests that the choice between Cas12a and Cas9 may depend on the specific editing strategy (single vs. multi-target).
In plant systems (tomato), LbCas12a and SpCas9 showed similar overall, though target-dependent, efficiencies. A notable difference was the mutational pattern: Cas12a induced more and larger deletions than Cas9, which can be advantageous for applications requiring complete gene disruption [4]. In maize, the CRISPR-Cas9 system showed higher editing efficiency in early generations (T0, T1), though Cas12a generated more diverse mutations in later generations (T2) [12].
Table 2: Comparative Performance of Cas12a and Cas9 in Experimental Systems
| Application / System | Cas12a Efficiency | Cas9 Efficiency | Key Finding |
|---|---|---|---|
| CAR-T Knock-in (Human CD8+ T-cells) | 37.4% - 51% (CLASH system) [25] | Not reported in this study | Cas12a enables massively parallel knock-in engineering. |
| Single-Gene Editing (A. niger) | 86.5% [23] | 31.7% [23] | Cas12a showed higher efficiency for single-target editing. |
| Dual-gRNA Editing (A. niger) | Up to 100% [23] | Up to 100% [23] | Both systems achieve maximal efficiency with multiple guides. |
| Gene Editing (Tomato Protoplasts) | Similar, but target-dependent [4] | Similar, but target-dependent [4] | Cas12a produced more and larger deletions. |
| Multiplex Editing (Maize) | Lower in T0/T1, more diverse mutations in T2 [12] | Higher in T0/T1 [12] | Performance varies across generations and outcome measures. |
Detailed Experimental Protocol: Cas12a-Mediated Knock-in in Primary Human T-Cells
The following protocol details the methodology for the CLASH system [25], which can be adapted for targeted knock-in in primary human T-cells.
CLASH Workflow for Massively Parallel CAR-T Engineering
Diagram 2: CLASH Workflow for Parallel CAR-T Engineering
1. CLASH Vector Design and Production
- Design the AAV Donor Vector: The AAV vector should contain three key components flanked by homology-directed repair (HDR) arms for the target locus (e.g., the TRAC locus for TCR knockout and CAR knock-in):
- CAR Transgene: Include an anti-tumor CAR construct (e.g., anti-CD22 CAR with scFv, transmembrane, and signaling domains).
- Targeting crRNA: Incorporate a constitutive crRNA targeting the first exon of the TRAC gene to facilitate knock-in.
- Wild-card crRNA Cassette: Design a pool of crRNAs targeting genes of interest (e.g., immune checkpoints, signaling molecules) for multiplexed knockout. Separate individual crRNAs within the array using Cas12a direct repeats (DRs) [25].
- Package the Vector: Package the final construct into AAV6 particles, which efficiently transduce primary human T-cells.
2. T-Cell Isolation and Activation
- Isolate primary human T-cells from healthy donor leukapheresis packs using density gradient centrifugation and negative selection kits.
- Activate T-cells using anti-CD3/CD28 magnetic beads and culture in appropriate medium supplemented with IL-2 for 2-3 days prior to editing [25] [27].
3. Delivery of CRISPR Components
- Electroporation: Deliver Cas12a (Cpf1) mRNA into activated T-cells via electroporation. Using mRNA rather than plasmid DNA reduces sustained Cas12a expression and minimizes cytotoxicity.
- Viral Transduction: Shortly after electroporation, transduce T-cells with the prepared pooled AAV6-CLASH library. AAV offers high transduction efficiency with low toxicity in T-cells [25].
4. Analysis and Validation
- At 5-7 days post-editing, analyze knock-in efficiency by flow cytometry, detecting TCR knockout (CD3-negative population) and successful CAR integration (CAR-positive population) [25].
- Perform targeted next-generation sequencing (NGS) on genomic DNA to verify on-target integration and assess the spectrum of induced mutations at the wild-card crRNA target sites.
- Subject the engineered T-cell pools to functional assays (e.g., cytokine release, cytotoxicity assays) and in vivo mouse models to select the most effective CAR-T variants [25].
The Scientist's Toolkit: Essential Reagents for Cas12a-Mediated Immune Cell Engineering
Table 3: Key Research Reagent Solutions for Cas12a T-Cell Engineering
| Reagent / Solution | Function | Examples / Notes |
|---|---|---|
| Cas12a Nuclease | Engineered variants for high fidelity and efficiency | LbCas12a, AsCas12a, enAsCas12a-HF1 [17] [25] |
| crRNA Library | Guides nuclease to specific genomic targets | Designed with 21-36 bp direct repeat; can be arrayed for multiplexing [25] [4] |
| AAV Donor Template | Provides HDR template for precise knock-in | AAV6 serotype; contains HDR arms, CAR transgene, crRNA cassette [25] |
| Electroporation System | Delivers CRISPR components into primary cells | Used for Cas12a mRNA delivery; optimized protocols for T-cell viability [25] [27] |
| T-Cell Activation Beads | Activates T-cells for efficient editing and proliferation | Anti-CD3/CD28 magnetic beads [27] |
| Cytokines | Supports T-cell growth and survival | IL-2 is commonly used to maintain T-cell cultures [27] |
Both Cas12a and Cas9 are highly effective tools for ex vivo immune cell engineering, yet they offer distinct advantages suited for different applications. Cas9 remains a robust choice for straightforward, high-efficiency single-gene knockout and has a proven track record in clinical development. In contrast, Cas12a excels in complex multiplexed editing scenarios, such as simultaneous knock-in of CAR transgenes and knockout of multiple endogenous genes, thanks to its simplified crRNA array processing and T-rich PAM preference. The development of advanced platforms like CLASH underscores Cas12a's unique utility for massively parallel engineering of primary human T-cells, enabling the unbiased discovery and selection of superior cell therapy products. The choice between these two powerful nucleases should be guided by the specific genetic engineering goals, target genomic loci, and the complexity of the desired immune cell phenotype.
The complexity of cancer, driven by combinations of multiple genetic mutations, has long posed a significant challenge for traditional disease modeling. The advent of CRISPR-based technologies, particularly when combined with the versatile delivery capabilities of adeno-associated virus (AAV), has revolutionized this field by enabling simultaneous perturbation of multiple genetic loci in living organisms (in vivo). This approach allows researchers to model the polygenic nature of cancer with unprecedented accuracy. Central to this revolution is the strategic choice of CRISPR effector—Cas9 or Cas12a—each offering distinct advantages for multiplexed genome editing in cancer induction studies. Cas9, the pioneering genome engineering workhorse, and Cas12a, a newer effector with unique capabilities for complex editing, now provide researchers with complementary tools for sophisticated in vivo cancer modeling [28] [2] [29].
This guide objectively compares the performance of AAV-delivered CRISPR-Cas9 and CRISPR-Cas12a systems for multiplexed cancer induction, providing experimental data, detailed methodologies, and practical resources to inform research design. We focus specifically on their application in creating autochthonous tumor models that recapitulate human disease progression within the native tissue microenvironment.
System Architectures: Cas9 and Cas12a Mechanisms
Fundamental Operational Differences
The architectural and mechanistic differences between Cas9 and Cas12a significantly influence their experimental applications for in vivo cancer modeling.
Cas9 System Architecture: The commonly used Streptococcus pyogenes Cas9 (SpCas9) requires two RNA components for operation: a CRISPR RNA (crRNA) containing the target-specific spacer sequence and a trans-activating crRNA (tracrRNA) that facilitates complex formation. These are often engineered as a single-guide RNA (sgRNA) for simplicity. Cas9 recognizes a 5'-NGG-3' protospacer adjacent motif (PAM) sequence and generates blunt-ended double-strand breaks (DSBs) in the target DNA [28] [29]. The blunt-end cuts are typically repaired by non-homologous end joining (NHEJ), often resulting in indel mutations that disrupt gene function—ideal for knocking out tumor suppressor genes in cancer models.
Cas12a System Architecture: Cas12a (formerly Cpf1) operates with a single crRNA and does not require a tracrRNA, simplifying vector design. It recognizes T-rich PAM sequences (5'-TTTV-3') and creates staggered DNA cuts with 5' overhangs [28] [2] [29]. These "sticky ends" can enhance homology-directed repair (HDR) efficiency in certain contexts and may promote more predictable repair outcomes. Cas12a's smaller size compared to SpCas9 is advantageous for AAV packaging, which has strict capacity limitations [28] [29].
Figure 1: Comparative workflows of AAV-delivered Cas9 and Cas12a systems for cancer induction. Cas9 requires two RNA components and creates blunt-end breaks, while Cas12a utilizes a single crRNA and generates staggered cuts with different PAM requirements.
crRNA Array Designs for Multiplexed Editing
A key advantage of Cas12a for complex cancer modeling is its innate capacity for processing multiple crRNAs from a single transcript. Cas12a can efficiently process a customized crRNA array, enabling simultaneous targeting of multiple oncogenes or tumor suppressor genes with a single vector [2] [30]. While Cas9 systems can also be engineered for multiplexing through tRNA or ribozyme-based processing systems, this requires additional synthetic biology components [23] [30].
For cancer induction, this multiplexing capability is crucial for modeling the cooperative effects of driver mutations. A single AAV vector can deliver a crRNA array targeting combinations like Trp53, Pten, Nf1, and Rb1—common alterations in glioblastoma, pancreatic cancer, and lung adenocarcinoma models [31] [30].
Quantitative Performance Comparison
Table 1: Direct comparison of Cas9 and Cas12a editing capabilities based on experimental data
| Parameter | CRISPR-Cas9 | CRISPR-Cas12a | Experimental Context |
|---|---|---|---|
| Editing Efficiency (Single Target) | 31.7% | 86.5% | Aspergillus niger albA locus [23] |
| Multiplex Editing Efficiency | Up to 100% with 2 gRNAs | Up to 100% with 2 gRNAs | Aspergillus niger [23] |
| Large Deletion Efficiency | Up to 69.1% for 40 kb deletions | Similar efficiency for large deletions | 3.5-40 kb regions in A. niger [23] |
| PAM Requirement | 5'-NGG-3' | 5'-TTTV-3' | Various species [28] [2] [29] |
| Cut Type | Blunt ends | Staggered cuts (5' overhangs) | In vitro characterization [28] [29] |
| RNA Components | gRNA + tracrRNA | Single crRNA | Various species [28] [2] [29] |
Table 2: In vivo tumor induction performance using AAV-CRISPR systems
| Parameter | AAV-Cas9 Library | Experimental Context |
|---|---|---|
| Tumor Incidence Rate | 50% (9/18 mice) at 4 months | LSL-Cas9 mice, LV/HPF injection [31] |
| Median Survival | 129 days (95% CI: 111-159 days) | LSL-Cas9 mice with AAV-mTSG library [31] |
| Tumor Volume | 70.2 mm³ (all mice), 140.3 mm³ (tumor-bearing only) | MRI measurement at 4 months [31] |
| Endpoint Tumor Prevalence | 91% (10/11 mice) | Histopathological confirmation [31] |
| Co-mutation Detection | Mll2, B2m-Nf1, Mll3-Nf1, Zc3h13-Rb1 | Capture sequencing analysis [31] |
Experimental Protocols for In Vivo Cancer Modeling
AAV-mediated Autochthonous CRISPR Screening for Glioblastoma
Background: This protocol, adapted from [31], enables functional interrogation of cancer genes through direct in vivo screening in immunocompetent mice. The approach models the natural evolution of tumors within their native microenvironment, capturing critical interactions between cancer cells and the immune system.
Materials and Methods:
- Animal Model: Conditional Rosa26-LSL-Cas9-GFP mice (LSL-Cas9 mice) [31]
- AAV Library: AAV-mTSG library targeting mouse homologs of pan-cancer significantly mutated genes, packaged into AAV vectors with GFP reporter and Cre recombinase under GFAP promoter [31]
- Controls: Empty AAV-vector and PBS-injected mice [31]
Procedure:
- Library Design: Pool-synthesize sgRNAs targeting cancer-related genes with >100× coverage; verify library representation by deep sequencing [31]
- Virus Production: Generate high-titer AAV preparations (>1×10¹² viral particles/mL) from the plasmid library [31]
- Stereotaxic Injection: Deliver AAV-mTSG library, AAV-vector control, or PBS into lateral ventricle (LV) or hippocampus (HPF) of LSL-Cas9 mice [31]
- Tumor Monitoring: Perform serial magnetic resonance imaging (MRI) at 4 months post-injection to detect tumor development and measure volumes [31]
- Survival Analysis: Monitor animals for survival endpoints (defined by poor body condition score BCS <2) over 299 days [31]
- Molecular Analysis: Harvest endpoint tumors for capture sequencing to identify mutational profiles and co-mutation patterns [31]
Key Modifications for Cas12a Implementation:
- Replace Cas9 sgRNAs with Cas12a crRNA arrays targeting the same gene sets
- Utilize Cas12a-expressing mouse models instead of Cas9 models
- Maintain identical AAV production and injection parameters for direct comparison
Cell-Type-Specific Screening Using CrAAVe-Seq
Background: The CRISPR screening by AAV episome-sequencing (CrAAVe-seq) platform enables cell-type-specific genetic screening in targeted populations within mouse tissues [32]. This technology is particularly valuable for studying cell-type-specific vulnerabilities in cancer initiation.
Materials and Methods:
- Vector Design: pAP215 AAV vector with Cre-sensitive sgRNA construct containing Lox71/Lox66-flanked handle cassette [32]
- Animal Model: Inducible CRISPR interference (Lox-Stop-Lox-dCas9-KRAB or LSL-CRISPRi) mice [32]
- AAV Capsid: PHP.eB capsid for widespread brain transduction [32]
Procedure:
- Virus Packaging: Package pAP215 sgRNA library into PHP.eB capsid [32]
- Co-injection: Deliver PHP.eB::pAP215 and cell-type-specific Cre recombinase (e.g., hSyn1-Cre for neurons) by intracerebroventricular (ICV) injection into neonatal mice [32]
- Episome Recovery: At endpoint, perform isopropanol precipitation of nucleic acids from TRIzol-chloroform extraction of whole mouse brain [32]
- Cell-Type-Specific Amplification: PCR amplify using primers specific to the Cre-inverted handle sequence to selectively quantify sgRNAs in Cre-expressing cells [32]
- Sequencing and Analysis: Sequence amplified products and analyze sgRNA frequencies to identify essential genes in specific cell populations [32]
Figure 2: Generalized workflow for AAV-CRISPR in vivo cancer induction. The process begins with library design and proceeds through viral production, precise delivery to target tissues, longitudinal monitoring, and final molecular analysis to characterize induced tumors.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key research reagents for implementing AAV-CRISPR cancer induction studies
| Reagent/Solution | Function | Examples/Specifications |
|---|---|---|
| Conditional Cas Mice | Provides tissue-specific Cas expression | Rosa26-LSL-Cas9-GFP [31], LSL-CRISPRi mice [32] |
| AAV Transfer Plasmids | sgRNA/crRNA expression for library cloning | pAP215 (Cre-dependent) [32], Custom crRNA array vectors [30] |
| AAV Capsids | Determines tropism and transduction efficiency | PHP.eB (enhanced CNS tropism) [32], Serotype-specific variants |
| Cell-Type-Specific Promoters | Restricts CRISPR activity to target cells | hSyn1 (neuronal) [32], GFAP (astrocytic) [31] |
| crRNA Array Processing Systems | Enables multiplexed targeting from single transcript | tRNA-gRNA arrays [23] [30], Cas12a innate processing [2] [30] |
| Sequencing Assays | Identifies mutational profiles and editing outcomes | Capture sequencing [31], AAV episome sequencing [32] |
| Viral Production Systems | Generates high-titer AAV preparations | HEK293 transfection, >1×10¹² vp/mL [31] |
Discussion: Strategic Implementation for Cancer Modeling
Context-Dependent System Selection
The choice between Cas9 and Cas12a for AAV-delivered cancer induction depends heavily on specific research goals and target genomic contexts. Cas9 remains the preferred option for researchers prioritizing established protocols, extensive validation data, and compatibility with existing mouse models. Its well-characterized behavior and the availability of numerous engineered variants provide a solid foundation for standardized cancer modeling approaches [31] [28].
Conversely, Cas12a offers distinct advantages for projects requiring enhanced multiplexing capacity, access to T-rich genomic regions, or more efficient AAV packaging due to its smaller size. The ability to process extended crRNA arrays from a single transcript makes Cas12a particularly valuable for modeling complex oncogenic collaborations involving 3+ genes [2] [30]. Furthermore, Cas12a's staggered cuts may promote more predictable repair outcomes in certain contexts, potentially reducing the screening burden for desired cancer models [2] [29].
Clinical Translation and Therapeutic Modeling
The progression of both Cas9 and Cas12a systems into clinical trials highlights their relevance for developing next-generation cancer therapies. Cas9-based approaches have demonstrated remarkable success in ex vivo applications, while Cas12a's compact size and multiplexing capabilities make it increasingly attractive for in vivo therapeutic genome editing [22] [33]. The recent development of Cas12a variants with enhanced editing efficiency, such as high-fidelity Cas12Max (hfCas12Max), further expands the toolkit for creating more precise cancer models that better recapitulate human disease genetics [33].
AAV-delivered CRISPR systems have fundamentally transformed in vivo cancer modeling by enabling sophisticated multiplexed genome editing in authentic tissue microenvironments. The strategic selection between Cas9 and Cas12a should be guided by specific experimental requirements: Cas9 offers proven reliability and extensive validation, while Cas12a provides superior multiplexing capacity and access to previously challenging genomic targets. As both systems continue to evolve through protein engineering and delivery optimization, their synergistic application will undoubtedly accelerate our understanding of cancer genetics and foster the development of increasingly precise cancer models that faithfully recapitulate human disease progression and therapeutic responses.
The ability to simultaneously perturb multiple genetic loci is crucial for deconvoluting complex biological phenomena, such as gene interaction networks, synthetic lethality, and multigenic diseases. While CRISPR-Cas9 has served as the foundational technology for genome engineering, the expanding CRISPR toolbox now includes Cas12a, which offers distinct advantages for multiplexed applications. Simultaneous Dual-Gene Activation and Knockout (DAKO) represents a cutting-edge advancement that enables researchers to activate and knock out genes concurrently within the same cell, providing a powerful system for modeling complex genetic interactions and screening for therapeutic targets [17]. This technology is particularly valuable for cancer modeling, immune cell engineering, and functional genomics studies where combinatorial gene perturbations are necessary to mimic disease states or elucidate functional pathways.
The development of DAKO systems leverages the unique molecular capabilities of CRISPR nucleases, especially Cas12a, which simplifies multiplexed editing through its inherent ability to process multiple guide RNAs from a single array. This article provides a comprehensive comparison of Cas9 and Cas12a platforms for multiplex editing research, with a specific focus on the implementation, efficiency, and applications of DAKO systems, supported by experimental data and detailed methodologies.
Molecular Mechanisms: Cas9 vs. Cas12a for Multiplexed Editing
Fundamental Biochemical Differences
Cas9 and Cas12a, both Class 2 CRISPR effectors, exhibit distinct structural and functional characteristics that influence their application in genome editing, particularly for multiplexed approaches like DAKO.
PAM Recognition and Targeting Space: Cas9 from Streptococcus pyogenes typically requires a 5'-NGG-3' protospacer adjacent motif (PAM) adjacent to its target sequence, which favors GC-rich genomic regions [2] [10]. In contrast, Cas12a recognizes a 5'-TTTV-3' PAM (where V is A, C, or G), making it particularly suitable for targeting AT-rich regions that are often challenging for Cas9 [2] [7]. This fundamental difference in PAM requirement expands the total targetable space in eukaryotic genomes when both systems are available.
DNA Cleavage Mechanism: A critical distinction lies in their DNA cleavage patterns. Cas9 generates blunt-ended double-strand breaks (DSBs) upstream of the PAM site, while Cas12a creates staggered cuts with 5' overhangs distal to the PAM recognition site [2] [18]. These staggered ends may facilitate more predictable repair outcomes and potentially enhance homology-directed repair (HDR) efficiency, which is advantageous for precise genome engineering applications [2].
Guide RNA Requirements: Cas9 requires two RNA components - a CRISPR RNA (crRNA) that contains the target-specific spacer and a trans-activating crRNA (tracrRNA) - which are often fused into a single-guide RNA (sgRNA) for simplicity [10]. Cas12a operates with only a single crRNA, lacking the need for a tracrRNA component. This simpler architecture is particularly beneficial for multiplexed editing, as it reduces the overall size of the genetic payload [10].
Multiplexing Capabilities
For simultaneous editing of multiple genomic loci, Cas12a offers distinct advantages due to its innate ability to process multiple crRNAs from a single transcript.
crRNA Array Processing: Cas12a possesses RNase activity that enables it to process a long crRNA array into individual mature crRNAs by cleaving at direct repeat (DR) sequences [17]. This autonomous processing capability allows for efficient multiplexed gene perturbation from a single transcriptional unit, simplifying the delivery construct design for targeting multiple genes simultaneously [17] [24].
Delivery Efficiency: The compact nature of Cas12a crRNAs (approximately 41 nucleotides) compared to Cas9 sgRNAs (approximately 100 nucleotides) reduces the size of the genetic payload, which is particularly advantageous for viral delivery systems with limited packaging capacity [17]. This feature enables the delivery of more complex multiplexed systems, including those required for DAKO applications.
Table 1: Fundamental Comparison of Cas9 and Cas12a Nucleases
| Characteristic | Cas9 | Cas12a |
|---|---|---|
| PAM Sequence | 5'-NGG-3' (G-rich) | 5'-TTTV-3' (T-rich) |
| Guide RNA | Dual RNA (crRNA + tracrRNA) or sgRNA | Single crRNA |
| crRNA Length | ~100 nt (sgRNA) | ~41 nt |
| DNA Cleavage | Blunt ends | Staggered cuts with 5' overhangs |
| Multiplexing | Requires multiple sgRNAs or complex processing systems | Innate crRNA array processing capability |
| Target Preference | GC-rich regions | AT-rich regions |
DAKO System Implementation: A Cas12a-Based Platform
System Architecture and Components
The DAKO system represents a sophisticated application of CRISPR technology that enables simultaneous activation of one gene while knocking out another in the same cell. The foundational implementation of DAKO, as described in recent literature, utilizes Cas12a-knock-in mice integrated with a CRISPR activation (CRISPRa) system [17].
The core components of this DAKO platform include:
Cas12a-Knock-In Mice: These transgenic mice feature conditional or constitutive expression of high-fidelity enhanced AsCas12a (enAsCas12a-HF1) or LbCas12a inserted at the Rosa26 locus. The expression is driven by a CAG promoter and controlled by a LoxP-3xPolyA-Stop-LoxP (LSL) cassette for tissue-specific regulation [17].
Nuclear Localization Signals (NLS): To optimize nuclear targeting and editing efficiency, the Cas12a constructs incorporate different NLS combinations. The LbCas12a includes SV40 NLS on the N-terminus and nucleoplasmin NLS on the C-terminus, while enAsCas12a features Egl-13 NLS on the N-terminus with an additional c-Myc NLS on the C-terminus [17].
Affinity Tags and Reporters: The system includes C-terminal 3xHA tags for LbCas12a and Myc tags for enAsCas12a, followed by a 2A self-cleavage peptide and enhanced GFP (eGFP) reporter. These elements facilitate the detection and tracking of Cas12a expression [17].
CRISPRa Integration: For the activation component, the system incorporates a dCas9-SPH CRISPR activation transgenic mouse line, which provides the necessary components for gene activation when combined with the Cas12a knockout capability [17].
Experimental Workflow and Validation
The implementation of DAKO follows a structured workflow that ensures precise targeting and validation of the dual-function outcomes:
Diagram Title: DAKO Experimental Workflow
The validation of DAKO systems includes comprehensive assessment at molecular and functional levels:
Molecular Validation: Confirmation of successful gene knockout through sequencing to detect indels at the target loci and verification of gene activation through mRNA expression analysis (e.g., RT-qPCR) and protein level assessment [17].
Functional Validation: Evaluation of phenotypic outcomes resulting from the combined genetic perturbations, which may include changes in cell differentiation, proliferation, or functional responses in specific cellular contexts [17].
Performance Comparison: Experimental Data
Editing Efficiency Across Platforms
Direct comparisons of Cas9 and Cas12a editing efficiencies have been conducted across various biological systems, providing valuable insights for researchers selecting appropriate platforms for multiplexed applications like DAKO.
Table 2: Comparative Editing Efficiencies of Cas9 and Cas12a Across Biological Systems
| Organism/System | Target | Cas9 Efficiency | Cas12a Efficiency | Notes | Citation |
|---|---|---|---|---|---|
| Chlamydomonas reinhardtii (green alga) | Multiple loci | ~20-30% total editing | Similar total editing (~20-30%), higher precision | Cas12a showed slightly higher precision in ssODN-templated editing | [13] |
| Rice (Oryza sativa) | Phytoene desaturase (PDS) | Lower efficiency | Higher efficiency (LbCas12a) | LbCas12a RNP achieved higher targeted mutagenesis frequency | [18] |
| Maize (Zea mays) | Glossy2 (gl2) gene | 90-100% of T0 plants had mutations | 0-60% of T0 plants had mutations | Cas9 significantly more efficient in this context | [10] |
| Aspergillus niger (fungus) | Polyketide synthase (albA) | 15% (single gRNA) | 89.5% (single crRNA) | Cas12a superior for single targeting | [34] |
| Aspergillus niger (fungus) | Multiple targets | Up to 100% (dual gRNAs) | Up to 100% (dual crRNAs) | Both systems highly efficient with multiple guides | [34] |
Specificity and Off-Target Profiles
Editing precision is a critical consideration, particularly for therapeutic applications. Comparative studies have revealed important differences in the off-target profiles of these two nucleases:
In Silico Off-Target Prediction: Analysis in maize revealed that Cas12a crRNAs have far fewer potential off-target sites in the genome compared to Cas9 gRNAs. One study reported 1,193 potential off-target sites for a Cas12a crRNA versus 19,029 for a Cas9 gRNA when allowing up to six mismatches [10].
Experimental Validation: Despite the in silico predictions, empirical validation in maize T1 plants revealed no detectable off-target mutations for Cas9 at the potential sites identified through CIRCLE-seq analysis [10]. This highlights the importance of experimental validation beyond computational predictions.
High-Fidelity Variants: Both Cas9 and Cas12a have been engineered for improved specificity. High-fidelity Cas9 variants (e.g., HiFi Cas9) demonstrate reduced off-target cleavage while maintaining on-target activity [18]. Similarly, enhanced Cas12a variants (e.g., enAsCas12a-HF1) have been developed with improved fidelity [17].
Research Reagent Solutions for DAKO Implementation
Successful implementation of DAKO systems requires specific reagents and tools. The following table outlines key components essential for establishing DAKO in a research setting.
Table 3: Essential Research Reagents for DAKO Systems
| Reagent/Tool | Function | Example/Specification | Application in DAKO | |
|---|---|---|---|---|
| Cas12a-Knock-In Mice | Provides constitutive or conditional Cas12a expression | LSL-enAsCas12a or LSL-LbCas12a at Rosa26 locus | Base platform for in vivo knockout component | [17] |
| dCas9-SPH Activator Line | Enables gene activation | Transgenic mouse line with dCas9 fused to activation domains | Activation component of DAKO system | [17] |
| crRNA Arrays | Targets Cas12a to specific genomic loci | Concatenated crRNAs with direct repeats for processing | Enables multiplexed knockout targeting | [17] |
| Viral Delivery Vectors | Delivers CRISPR components to cells | AAV, lentiviral, or retroviral vectors | Efficient delivery of crRNAs and activation components | [17] |
| Non-Viral Delivery Systems | Alternative delivery method | Lipid nanoparticles (LNPs) with synthetic crRNAs | In vivo delivery of editing components | [17] |
| Flex-Cas12a Variants | Expanded PAM recognition | Engineered LbCas12a with 6 mutations (e.g., G146R, R182V) | Increases targetable genomic space | [7] |
Applications in Biomedical Research
Disease Modeling and Functional Genomics
The DAKO system enables sophisticated disease modeling that more accurately reflects the complexity of human pathologies:
Cancer Modeling: DAKO facilitates the study of oncogene activation simultaneous with tumor suppressor knockout, better recapitulating the multistep process of carcinogenesis. Research has demonstrated efficient quadruplex gene knockout in vivo using a single AAV vector simultaneously targeting murine Trp53, Apc, Pten, and Rb1, resulting in rapid induction of salivary gland squamous cell carcinoma and lung adenocarcinoma [17].
Immune Cell Engineering: The system has been applied to retrovirus-based engineering of CD4+ and CD8+ T cells, B cells, and bone-marrow-derived dendritic cells, enabling simultaneous knockout of endogenous receptors while activating beneficial genes for enhanced function [17].
Functional Genomics: DAKO enables the study of genetic interactions, such as epistasis, redundancy, synergy, and antagonism, by allowing simultaneous perturbation of multiple genes in a single experiment [17] [24].
Therapeutic Development
The precision and multiplexing capabilities of Cas12a-based systems offer significant promise for therapeutic applications:
Gene Therapy: The ability to perform multiple genetic modifications simultaneously is valuable for complex gene therapy approaches, such as correcting mutations while simultaneously modulating expression of modifier genes [2].
Cell-Based Therapies: DAKO systems can enhance engineered cell therapies (e.g., CAR-T cells) by simultaneously knocking out inhibitory receptors while activating beneficial pathways to enhance persistence and efficacy [17].
Technical Considerations and Protocol Optimization
Critical Experimental Parameters
Successful implementation of DAKO requires optimization of several key parameters:
Delivery Method Selection: The choice of delivery vector significantly impacts editing efficiency. Viral vectors (AAV, lentivirus) offer high transduction efficiency, while lipid nanoparticles provide transient delivery with reduced immunogenicity concerns [17].
crRNA Array Design: For multiplexed Cas12a editing, crRNA arrays should be designed with appropriate direct repeats between spacer sequences to ensure efficient processing. The length and secondary structure of the array can impact processing efficiency [17].
Expression Level Optimization: Constitutive expression of Cas12a in knock-in mice did not lead to discernible pathology, indicating good tolerability [17]. However, in other systems, expression levels may need optimization to balance efficiency and potential cytotoxicity.
Troubleshooting Common Challenges
Low Knockout Efficiency: If knockout efficiency is suboptimal, consider testing alternative crRNA sequences, verifying Cas12a nuclear localization, or using high-fidelity variants (enAsCas12a-HF1) with expanded PAM recognition [17] [7].
Inadequate Activation: For the activation component, ensure proper delivery and expression of the dCas9-SPH system and verify guide RNA design for activation elements targeting promoter regions [17].
Cellular Toxicity: Monitor for potential toxicity from multiple DSBs or high levels of CRISPR component expression. Implement strategies for transient expression or titrate delivery to minimize stress while maintaining editing efficiency [24].
Future Directions and Engineering Advances
Recent protein engineering efforts have addressed some limitations of native Cas12a systems, particularly expanding the targeting range:
PAM Relaxation: Directed evolution approaches have generated Cas12a variants with expanded PAM recognition. The Flex-Cas12a variant, carrying six mutations (G146R, R182V, D535G, S551F, D665N, and E795Q), recognizes 5'-NYHV-3' PAMs, expanding potential genome accessibility from ~1% to over 25% [7].
Enhanced Specificity: Continued engineering focuses on reducing off-target effects while maintaining high on-target activity, through both structure-guided mutations and directed evolution approaches [18] [7].
Novel Delivery Strategies: Advances in delivery modalities, including improved viral vectors and non-viral nanoparticles, are enhancing the efficiency and specificity of DAKO component delivery to target tissues [17].
The development of DAKO systems represents a significant advancement in multiplex genome engineering, providing researchers with a powerful tool to model complex genetic interactions and develop novel therapeutic strategies. As Cas12a engineering continues to evolve, these systems will likely become more efficient, specific, and accessible to the research community.
Enhancing Efficiency and Precision in Multiplex CRISPR Workflows
A key challenge in multiplexed genome engineering is the inconsistent performance of individual crRNAs, which can compromise the overall success of an experiment. Both the choice of CRISPR system (Cas9 vs. Cas12a) and specific optimization strategies significantly influence editing outcomes. The table below summarizes quantitative data on strategies for improving editing efficiency.
| Strategy | Experimental System | Key Efficiency Metric | Reported Outcome |
|---|---|---|---|
| Cas12a for Multiplexed Base Editing | Human cells (HEK293) [35] | Editing frequency at 15 target sites | Achieved multiplexed editing at 15 loci, a 3-fold increase over previous state-of-the-art [35]. |
| Exonuclease Fusion (sbcB-LbCas12a) | Rice protoplasts [36] | Proportion of deletions >15 bp | Induced a 3.6-fold average increase in large deletions compared to LbCas12a alone [36]. |
| Multiplex gRNA Vector | Arabidopsis T1 plants [37] | Mutagenesis frequency at 6 target genes | Achieved mutations in all six targeted genes in one line; individual target mutation rates ranged from 13% to 93% [37]. |
| gRNA Array GC Content Optimization | Human cells with LbABE8e [35] | Editing frequency with different gRNA partners | Editing efficiency was significantly influenced by the %GC content and position of adjacent gRNAs in an array [35]. |
| Cas9 vs. Cas12a RNP Delivery | Chlamydomonas reinhardtii [13] | Total editing levels & precision | Both induced 20-30% editing; Cas12a showed slightly higher precision in ssODN-templated editing [13]. |
Nuclease Selection: Cas9 vs. Cas12a for Multiplexing
The fundamental choice between Cas9 and Cas12a is crucial, as each offers distinct advantages for multiplexing applications.
- Locus Targeting Capacity: Cas9 holds a practical advantage in the sheer number of sites it can target. Analyses show Cas9 can access 8 times more target sites within promoter regions and 32 times more within coding sequences compared to Cas12a [13].
- Multiplexed gRNA Expression: Cas12a has a unique biochemical advantage. Unlike Cas9, the Cas12a nuclease itself can process a single gRNA array transcript into multiple individual gRNAs without the need for additional accessory factors or complex promoter arrangements [35]. This simplifies vector design, reduces repetitive sequences that can cause genetic instability, and facilitates more efficient multiplexing [35].
- Editing Precision and Outcomes: Cas12a demonstrates slightly higher precision when using single-stranded oligodeoxynucleotide (ssODN) repair templates [13]. Furthermore, Cas12a generates staggered-ended DNA double-strand breaks, which typically lead to larger deletions (6-14 bp) compared to the blunt ends and smaller indels created by Cas9 [36].
gRNA Engineering and Design Optimization
Optimizing the guide RNA itself is one of the most direct ways to rescue low efficiency.
- gRNA Design Parameters: For any Cas nuclease, the gRNA sequence must be unique to the target and possess a GC content between 40% and 80% for optimal stability and activity [38]. The target sequence should be checked for potential off-target sites using specialized software [38].
- gRNA Array Engineering for Cas12a: When designing gRNA arrays for Cas12a, the sequence context matters. The GC content of adjacent gRNAs can influence the secondary structure of the entire array transcript, which in turn affects how efficiently Cas12a processes it and the resulting editing efficiency at each target site [35]. Empirical testing of different gRNA combinations and positions within the array may be necessary for optimal results.
- Truncated gRNAs for Reduced Bystander Mutations: In base editing applications, a key challenge is "bystander mutations," where the deaminase enzyme modifies non-target bases nearby. Research shows that truncating the length of Cas12a gRNAs can help direct editing outcomes toward a single desired base-pair conversion, thereby increasing precision and reducing bystander effects [35].
Advanced Molecular Tools and Delivery Methods
Beyond guide design, leveraging advanced editor architectures and delivery formats can enhance performance.
- Exonuclease Fusions for Larger Deletions: Fusing exonucleases to Cas nucleases is a powerful strategy to alter editing profiles. For example, fusing the E. coli exonuclease sbcB to LbCas12a shifted the indel spectrum toward significantly larger deletions, outperforming other exonuclease fusions like TREX2 and T5 [36]. This is particularly useful for knocking out non-coding RNA genes or regulatory elements.
- Delivery as Ribonucleoprotein (RNP): Delivering pre-assembled complexes of Cas protein and gRNA as RNPs can accelerate editing and reduce off-target effects. Studies comparing Cas9 and Cas12a RNPs have shown that both can achieve high total editing levels (20-30%), with Cas12a offering a potential advantage in precision [13].
- High-Fidelity Cas Variants: For both Cas9 and Cas12a, engineered high-fidelity versions are available. These variants, such as eSpCas9(1.1) or HypaCas9, contain mutations that reduce off-target activity by weakening non-specific interactions with the DNA backbone or enhancing the enzyme's proofreading capability [39].
Experimental Protocols for Key Optimization Strategies
Protocol 1: Testing Cas12a gRNA Array Performance
This protocol is adapted from studies demonstrating highly multiplexed base editing in human cells [35].
- gRNA Array Cloning: Clone a polycistronic gRNA array, containing up to 15 target sequences, into a plasmid under the control of the human U6 (hU6) promoter. The array relies on the native processing activity of Cas12a.
- Plasmid Transfection: Co-transfect the gRNA array plasmid with a plasmid expressing a Cas12a-derived base editor (e.g., LbCas12a-ABE or CBE) into HEK293 cells.
- Selection and Outgrowth: Treat transfected cells with puromycin (2 µg/mL) for selection. Allow for an outgrowth phase of 7 days post-transfection to ensure robust editing.
- Efficiency Assessment: Harvest genomic DNA and analyze editing efficiency at each target locus within the array using next-generation sequencing (NGS). This allows for the quantification of editing frequency and the identification of any performance variations between individual gRNAs.
Protocol 2: Enhancing Deletion Sizes with Exonuclease Fusions
This protocol is based on work in rice protoplasts to increase Cas12a-mediated deletion sizes [36].
- Construct Fusion Editor: Genetically fuse the exonuclease sbcB to the N-terminus of LbCas12a using a flexible linker (e.g., XTEN).
- Delivery to Protoplasts: Introduce the constructed sbcB-LbCas12a fusion vector, along with a multiplexed gRNA construct targeting multiple independent genomic sites, into rice protoplasts using a standard transfection method.
- Sequencing and Analysis: After 48-72 hours, extract DNA and perform targeted deep sequencing of the loci. Analyze the resulting indel spectra, specifically calculating the proportion of deletions exceeding 15 bp compared to controls with LbCas12a alone.
Workflow for Optimizing Low-Performing crRNAs
The following diagram illustrates a logical, step-by-step workflow for diagnosing and addressing the issue of a low-performing crRNA.
Research Reagent Solutions
The table below lists key reagents and their functions for implementing the strategies discussed in this guide.
| Reagent / Tool | Function in Optimization | Application Context |
|---|---|---|
| LbCas12a-derived Base Editor (e.g., LbABE8e) | Enables precise A-to-G base conversions and simplified multiplexing via gRNA array processing [35]. | Multiplexed base editing in human cells. |
| sbcB-LbCas12a Fusion Protein | Increases the size of genomic deletions by fusing an exonuclease to the nuclease, promoting larger deletions [36]. | Gene knockout, regulatory element disruption in plants. |
| Synthetic sgRNA | Provides high-purity, chemically synthesized guides for consistent performance and reduced off-target effects compared to plasmid-based expression [38]. | High-efficiency editing in all systems, especially with RNP delivery. |
| Multiplex gRNA Vector (Golden Gate Assembly) | Allows modular assembly of multiple gRNA expression cassettes with different promoters into a single vector to avoid recombination [37] [24]. | Creating higher-order combinatorial knockouts in plants and animals. |
| High-Fidelity Cas9 (e.g., SpCas9-HF1) | Reduces off-target editing by incorporating mutations that disrupt non-specific interactions with the DNA phosphate backbone [39]. | Experiments where minimizing off-target effects is critical. |
| TREX2-SpCas9 Fusion | Increases the frequency of large deletions and microhomology-mediated end joining (MMEJ) when using SpCas9 [36]. | Inducing larger deletions with Cas9 for gene or regulatory element knockout. |
| U6 / 7SL RNA Pol III Promoters | Drive high-level expression of gRNAs; using different promoters for different gRNAs in a multiplex vector prevents homologous recombination [37]. | Constructing stable multiplex vectors for plant and mammalian cells. |
The CRISPR-Cas gene-editing landscape has been predominantly shaped by the Cas9 system. However, for complex research applications requiring the simultaneous modification of multiple genes, known as multiplex editing, Cas9 faces inherent limitations. The recent development of hyper-efficient Cas12a variants represents a significant protein engineering breakthrough, offering a powerful alternative that excels in multiplexing scenarios. This guide provides an objective comparison of the performance of these novel Cas12a variants against other CRISPR systems, including canonical Cas9 and wild-type Cas12a, to inform researchers and drug development professionals.
Cas12a vs. Cas9: A Fundamental Comparison for Editing
Cas12a and Cas9, both Class II CRISPR-associated nucleases, differ in several key aspects that influence their application in research and therapy. Understanding these fundamental differences is crucial for selecting the right tool for your experiments.
The table below summarizes the core distinctions:
Table: Fundamental Differences Between Cas9 and Cas12a Systems
| Feature | Cas9 System | Cas12a System |
|---|---|---|
| PAM Sequence | 3'-NGG-3' [2] [40] | 5'-TTTV-3' (Wild-type & V3) [40]; 5'-TTTN* (Ultra) [40] |
| Guide RNA | Two-part (crRNA & tracrRNA) or single-guide RNA (sgRNA) [2] [40] | Single crRNA (42-44 nt) [2] [40] |
| DNA Cleavage | Blunt-ended double-strand breaks [2] | Staggered cuts with 5' overhangs [2] [40] |
| Native Multiplexing | Requires additional machinery for processing multiple gRNAs [35] | Can naturally process its own CRISPR arrays [35] [4] |
| Ideal for Genomes/Regions | General purpose; GG-rich regions [2] [40] | AT-rich genomes; regions with limited Cas9 design space [2] [40] |
Note: The Cas12a Ultra variant recognizes a TTTN PAM, which is even more relaxed than the canonical TTTV (V = A, C, or G) [40].
These inherent characteristics give Cas12a distinct advantages for multiplexed genome editing. Its ability to process a single RNA transcript into multiple crRNAs using its intrinsic RNase activity simplifies the delivery of complex editing systems [35]. Furthermore, the staggered DNA ends it creates may be more favorable for certain DNA repair pathways, potentially leading to more predictable editing outcomes [2].
The Emergence of Hyper-Efficient Cas12a Variants
The primary limitation of wild-type Cas12a has been its restrictive PAM requirement (TTTV), which confines its targeting scope to only about ~1% of a typical genome [7]. To overcome this, scientists have employed advanced protein engineering strategies, most notably directed evolution.
Flex-Cas12a: A Breakthrough in PAM Recognition
One of the most significant breakthroughs is the development of Flex-Cas12a. Using a bacterial-based directed evolution assay combined with rational engineering, researchers identified a variant of Lachnospiraceae bacterium Cas12a (LbCas12a) carrying six mutations (G146R, R182V, D535G, S551F, D665N, and E795Q) [7]. This variant exhibits dramatically expanded PAM recognition.
- Expanded Targeting Scope: Flex-Cas12a utilizes 5'-NYHV-3' PAMs (where N is any base, Y is C or T, and H is A, C, or T), increasing the proportion of targetable sites in the human genome from approximately 1% to over 25% [7].
- Retained Canonical Activity: A key advantage of Flex-Cas12a is that it retains efficient cleavage activity at the original 5'-TTTV-3' PAM sites, making it a versatile drop-in replacement for existing Cas12a workflows [7].
Cas12a Ultra: Commercial Application
The commercial sector has also advanced Cas12a engineering. The Alt-R Cas12a (Cpf1) Ultra is described as recognizing a broad TTTN PAM, further increasing the number of available target sites for researchers [40].
Performance Comparison: Experimental Data
The following tables consolidate key experimental data from recent studies, comparing the performance of engineered Cas12a variants against wild-type Cas12a and Cas9.
Table 1: Comparison of Editing Efficiency and Specificity
| System | Editing Efficiency | Mutation Pattern | Off-Target Notes |
|---|---|---|---|
| SpCas9 | Target-dependent; shown to be efficient in tomato [4] | Predominantly small insertions/deletions (indels); blunt ends [4] | Well-characterized; off-target sites can be predicted |
| LbCas12a (Wild-type) | Similar overall efficiency to SpCas9, though target-dependent [4] | More and larger deletions than Cas9; staggered ends [4] | Off-target activity found with 1-2 mismatches distal from PAM; generally precise [4] |
| Flex-Cas12a | Retained robust nuclease activity at canonical and non-canonical PAMs [7] | Retains wild-type cleavage kinetics, reducing promiscuous targeting [7] | Designed to maintain high specificity despite relaxed PAM [7] |
Table 2: Comparison of Multiplex Editing Capabilities
| System | Multiplexing Efficiency | Key Experimental Findings |
|---|---|---|
| Cas9-derived Base Editors | Limited by complex gRNA delivery; repetitive sequences cause instability [35] | Lower efficiency in simultaneous editing of multiple loci [35] |
| dLbCas12a-derived Base Editors | Highly effective for multiplexed base editing (MBE) [35] | Capable of editing up to 15 endogenous target sites simultaneously in human cell lines [35] |
| Cas12a (LbCas12a) | Robust deletion efficiency in large genomic fragments [41] | Achieved up to 100% editing efficiency for single gene edits in Aspergillus niger; successfully deleted fragments over 100 kb [41] |
Experimental Protocols for Key Studies
Directed Evolution of Flex-Cas12a
The following diagram illustrates the workflow used to develop the hyper-efficient Flex-Cas12a variant.
Detailed Methodology [7]:
- Library Generation: Researchers performed error-prone PCR on the DNA fragment encoding the PAM-interacting (PI) and wedge (WED) domains of LbCas12a to introduce random mutations.
- Selection System: The mutant libraries were subjected to a dual-bacterial selection system. Cas12a variants were co-expressed with a crRNA targeting a sequence next to a non-canonical PAM (e.g., AGCT, AGTC) within a lethal ccdB gene on a plasmid.
- Enrichment: Only Cas12a variants capable of cleaving the lethal gene at the non-canonical PAM site allowed bacterial survival on selective media, enabling the isolation of PAM-relaxed variants.
- Validation: Positive clones were sequenced, and the leading candidate, Flex-Cas12a, was characterized using in vitro biochemical assays and editing assays in human cells to confirm PAM preference and editing efficiency.
Evaluating Multiplex Base Editing with Cas12a
The following diagram outlines the process for testing high-order multiplex base editing.
Detailed Methodology [35]:
- Base Editor Construction: Catalytically dead LbCas12a (dLbCas12a) was fused to deaminase moieties (e.g., APOBEC3A for CBE, ABE8e for ABE) to create Cas12a-derived base editors.
- gRNA Array Cloning: Multiple individual gRNAs targeting genes like RUNX1, DNMT1, and EMX1 were cloned into a single expression cassette. Cas12a's native ability to process its own arrays was leveraged to generate individual crRNAs from a single transcript.
- Cell Transfection & Culture: The BE and gRNA array plasmids were co-transfected into HEK293 cells. A critical optimization was the use of 2 µg/mL puromycin selection followed by a 7-day outgrowth phase to enrich for successfully edited cells.
- Outcome Measurement: Editing efficiency and the rate of bystander mutations (unintended edits near the target) were quantified at each target site using next-generation sequencing (NGS) of amplified genomic regions.
The Scientist's Toolkit: Essential Research Reagents
To replicate or build upon these studies, researchers will require the following key reagents:
Table 3: Essential Research Reagents for Cas12a Engineering and Application
| Reagent / Tool | Function & Importance |
|---|---|
| LbCas12a Expression Plasmid | Source of the Cas12a nuclease. For hyper-efficient variants, the plasmid must encode the specific mutations (e.g., Flex-Cas12a mutations) [7]. |
| crRNA Expression Vector | A plasmid or kit system for cloning and expressing single crRNAs or crRNA arrays. Golden Gate assembly-compatible systems are widely used [4]. |
| Directed Evolution Selection System | A bacterial system (e.g., with a ccdB negative selection gene) for screening and isolating Cas12a variants with desired PAM relaxation [7]. |
| Cell Lines for Validation | Standard mammalian cell lines (e.g., HEK293) or other relevant cell types (e.g., tomato protoplasts, Aspergillus niger) for testing editing efficiency and specificity [7] [4] [41]. |
| NGS Amplicon-Seq Kit | Essential for high-throughput, quantitative analysis of on-target editing efficiency and the detection of off-target effects [4]. |
The protein engineering of hyper-efficient Cas12a variants like Flex-Cas12a marks a pivotal advancement in the CRISPR toolkit. For researchers focused on multiplex editing, these variants offer a compelling combination of a dramatically expanded targeting scope (≥25% of the genome), high efficiency in multi-gene knockouts and base editing (up to 15 sites), and the inherent advantages of the Cas12a system, such as simplified guide RNA design and staggered DNA breaks. While Cas9 remains a powerful and versatile tool, engineered Cas12a variants now provide a superior alternative for complex genetic engineering tasks in both basic research and therapeutic development.
The advent of CRISPR-Cas systems has revolutionized genetic engineering, offering researchers unprecedented control over genome manipulation. Among the various CRISPR nucleases available, Cas9 and Cas12a have emerged as two of the most powerful and widely used tools for targeted genome editing. While both systems function as RNA-guided endonucleases that create double-strand breaks in DNA, they differ significantly in their mechanisms, specificities, and optimal applications. For researchers, scientists, and drug development professionals, understanding the off-target profiles of these systems is paramount for experimental design and therapeutic development. This guide provides a comprehensive, evidence-based comparison of Cas9 and Cas12a specificity, drawing from recent scientific investigations across various biological systems to inform your genome editing strategies.
Fundamental Structural and Mechanistic Differences
Cas9 and Cas12a originate from distinct evolutionary pathways, resulting in different molecular architectures and cleavage mechanisms that fundamentally influence their editing specificities.
CRISPR-Cas9, particularly from Streptococcus pyogenes (SpCas9), requires two RNA components: a CRISPR RNA (crRNA) that specifies the target sequence and a trans-activating crRNA (tracrRNA) that facilitates complex formation. These are often combined into a single guide RNA (sgRNA) for experimental applications [2]. Cas9 recognizes G-rich protospacer adjacent motifs (PAMs) with the sequence 5'-NGG-3' located immediately downstream of the target site [42]. Upon target recognition, Cas9 generates blunt-ended double-strand breaks typically between the 17th and 18th nucleotides upstream of the PAM site [42].
CRISPR-Cas12a (also known as Cpf1) represents a more recently discovered system with several distinct characteristics. Unlike Cas9, Cas12a requires only a single CRISPR RNA (crRNA) and does not need a tracrRNA [10] [12]. It recognizes T-rich PAM sequences (5'-TTTV-3') located upstream of the target site [2] [42]. Cas12a creates staggered DNA cuts with 4-5 nucleotide 5' overhangs [42], which some studies suggest may promote more predictable repair outcomes through homology-directed repair pathways [2].
Table 1: Fundamental Characteristics of Cas9 and Cas12a Nucleases
| Characteristic | Cas9 | Cas12a |
|---|---|---|
| PAM Sequence | 5'-NGG-3' (downstream) | 5'-TTTV-3' (upstream) |
| Guide RNA | Dual RNA (crRNA + tracrRNA) or sgRNA | Single crRNA |
| Cleavage Pattern | Blunt ends | Staggered cuts with 5' overhangs |
| crRNA Processing | Requires host RNases or engineered gRNAs | Native RNase activity processes pre-crRNA arrays |
| Size | ~1000-1600 amino acids | ~1100-1300 amino acids |
Experimental Evidence: Direct Comparisons of Editing Specificity
Maize Transformation Studies
A comprehensive 2018 study directly compared Cas9 and Cas12a activities by targeting the maize glossy2 gene coding region with overlapping sequences recognized by both nucleases [10]. The researchers introduced constructs carrying SpCas9-gRNA and LbCas12a-crRNA into maize inbred B104 embryos using Agrobacterium-mediated transformation.
On-target mutation analysis revealed striking differences: 90%-100% of Cas9-edited T0 plants carried indel mutations, with 63%-77% being homozygous or biallelic mutants [10]. In contrast, only 0%-60% of Cas12a-edited T0 plants showed on-target mutations, indicating significantly lower editing efficiency under the experimental conditions used [10].
The study employed CIRCLE-seq analysis to identify genome-wide potential off-target sites for Cas9, finding 18 and 67 potential off-targets for the two gRNAs tested, with an average of five mismatches compared to the target sites [10]. Importantly, sequencing analysis of selected off-target sites in T1 plants revealed no detectable mutations at these sites, suggesting that despite the theoretical potential, actual off-target effects in stable transgenic maize plants were minimal [10].
Tomato Protoplast Investigations
A 2024 study provided additional insights through high-throughput amplicon sequencing of tomato protoplasts, comparing LbCas12a and SpCas9 at 35 overlapping target sites [4]. The research found that LbCas12a demonstrated similar overall efficiency to SpCas9, though this was highly target-dependent [4].
Notably, the mutational patterns differed significantly: Cas12a induced more and larger deletions than Cas9, a characteristic that could be advantageous for specific genome editing applications where complete gene disruption is desired [4]. Off-target activity for LbCas12a was detected at 10 out of 57 investigated sites, with one or two mismatches present distal from the PAM in all cases [4]. This systematic comparison concluded that Cas12a-mediated genome editing is generally precise when potential off-target sites with distal mismatches are avoided during target selection [4].
Multiplex Editing Capabilities in Maize
A 2021 investigation compared CRISPR/Cpf1 and CRISPR/Cas9 systems for multiplex gene editing in maize, targeting the bZIP transcription factor Opaque2 (O2) [12]. The study found that in T0 and T1 generations, the CRISPR/Cpf1 system showed lower editing efficiency than CRISPR/Cas9 [12]. However, in the T2 generation, CRISPR/Cpf1 generated more types of new mutations, suggesting different long-term editing dynamics [12].
While the CRISPR/Cas9 system tended to edit within the on-target range, the CRISPR/Cpf1 system preferred to edit between targets [12]. The editing efficiency in the CRISPR/Cpf1 system positively correlated with Cpf1 expression level, highlighting the importance of promoter selection and expression optimization for achieving desired editing outcomes [12].
Table 2: Comparative Editing Efficiencies Across Experimental Systems
| Organism/System | Cas9 Efficiency | Cas12a Efficiency | Key Findings |
|---|---|---|---|
| Maize (T0 plants) | 90%-100% mutation rate [10] | 0%-60% mutation rate [10] | Cas9 more efficient in initial transformation |
| Tomato Protoplasts | Target-dependent efficiency [4] | Similar overall efficiency, target-dependent [4] | Cas12a produces larger deletions |
| Maize Multiplex Editing | Higher efficiency in T0-T1 [12] | Lower efficiency in T0-T1, diverse mutations in T2 [12] | Generational differences in mutation patterns |
| Aspergillus niger | 31.7% efficiency (single gRNA) [41] | 86.5% efficiency (single crRNA) [41] | Context-dependent efficiency variations |
Methodologies for Assessing Off-Target Effects
Computational Prediction Tools
Several in silico approaches have been developed to predict potential off-target sites, each with distinct algorithms and applications:
- Cas-OFFinder: Allows customizable parameters including sgRNA length, PAM type, and number of mismatches or bulges [43]. This tool identified far fewer potential off-target sites for Cas12a crRNAs compared to Cas9 gRNAs in maize [10].
- CCTop (Consensus Constrained TOPology prediction): Utilizes a scoring model based on the distances of mismatches to the PAM sequence [43].
- CFD (Cutting Frequency Determination): Employs experimentally validated datasets to predict cleavage likelihood [43].
- DeepCRISPR: Incorporates both sequence and epigenetic features in its predictions [43].
Experimental Detection Methods
Various experimental approaches have been developed to empirically identify off-target effects:
- CIRCLE-seq: A highly sensitive in vitro method that circularizes sheared genomic DNA, incubates it with Cas9/gRNA ribonucleoprotein complexes, then sequences the linearized DNA [10] [43]. This method identified 18 and 67 potential off-target sites for two Cas9 gRNAs in maize [10].
- GUIDE-seq: Utilizes double-stranded oligodeoxynucleotides (dsODNs) that integrate into double-strand breaks, enabling genome-wide identification of off-target sites with high sensitivity and low false-positive rates [43].
- Digenome-seq: Digests purified genomic DNA with Cas9/gRNA RNP complexes followed by whole-genome sequencing, offering high sensitivity but requiring substantial sequencing depth [43].
- Discover-seq: Leverages the DNA repair protein MRE11 as bait to perform chromatin immunoprecipitation followed by sequencing, providing high sensitivity and precision in cellular contexts [43].
Strategic Approaches for Minimizing Off-Target Effects
Target Selection Optimization
Careful target selection is the first line of defense against off-target effects:
- PAM Specificity: Cas12a's T-rich PAM (TTTV) provides access to genomic regions difficult to target with Cas9, potentially offering alternative editing sites with better specificity profiles [2].
- Bioinformatic Screening: Comprehensive genome-wide screening for sequences with high similarity to intended targets can identify potential off-target sites before experimental implementation [10] [4].
- Specificity Scoring: Utilizing tools that incorporate mismatch position effects (e.g., mismatches distal to PAM are generally better tolerated) can help select optimal targets [43] [4].
Experimental Design Considerations
- RNP Delivery: Using preassembled ribonucleoprotein (RNP) complexes rather than plasmid-based expression can limit the temporal window of nuclease activity, potentially reducing off-target effects [43].
- Modified Guide RNAs: Incorporating chemical modifications or structural alterations to guide RNAs can enhance specificity by increasing the energy requirement for off-target binding [43].
- High-Fidelity Variants: Engineered Cas9 and Cas12a variants with enhanced specificity profiles are increasingly available, though often with trade-offs in on-target efficiency [43].
Table 3: Key Research Reagents for Specificity-Focused Genome Editing
| Reagent/Resource | Function | Application Notes |
|---|---|---|
| Cas-OFFinder | Identifies potential off-target sites | Customizable PAM and mismatch parameters [43] |
| CIRCLE-seq | Genome-wide off-target detection | High sensitivity, in vitro method [10] [43] |
| GUIDE-seq | In-cell off-target identification | Uses dsODN integration into DSBs [43] |
| LbCas12a Expression Vector | Cas12a nuclease delivery | Codon optimization improves efficiency in plants [12] |
| crRNA Expression Cassettes | Guide RNA delivery | Promoter selection affects efficiency [4] |
| High-Fidelity Cas Variants | Enhanced specificity nucleases | Often trade reduced off-targets for lower on-target efficiency [43] |
The comparative analysis of Cas9 and Cas12a specificity reveals a complex landscape where multiple factors influence off-target effects. Current evidence suggests that both systems can achieve high specificity with careful experimental design, though their performance characteristics differ significantly across biological contexts.
Cas9 generally demonstrates higher editing efficiency in most systems studied, particularly in plants [10] [12], while Cas12a offers advantages through its distinct PAM requirements that expand targetable genomic regions [2] and its tendency to produce larger deletions that may be preferable for certain applications [4]. The native multiplexing capability of Cas12a through crRNA arrays presents another consideration for complex editing projects [12].
For researchers prioritizing minimal off-target effects, the current evidence supports:
- Comprehensive bioinformatic prediction during target selection
- Empirical validation using sensitive detection methods like CIRCLE-seq or GUIDE-seq
- Context-specific testing as nuclease performance varies across cell types and organisms
- Consideration of high-fidelity variants when maximal specificity is required
As CRISPR technologies continue to evolve, the development of next-generation nucleases with enhanced specificity profiles and improved prediction tools will further empower researchers to minimize off-target effects while maintaining high on-target activity.
The selection of a CRISPR-Cas system is a critical determinant in the success of any gene-editing experiment, particularly when the workflow involves delivery via viral vectors or lipid nanoparticles (LNPs). A primary challenge in the field is the fundamental trade-off between the robust editing capabilities of CRISPR nucleases and their physical size, which can exceed the packaging capacity of common delivery vehicles. This guide provides an objective comparison of Cas9 and Cas12a, focusing on how their distinct molecular characteristics—especially their size—impact practical delivery and application in multiplex editing research for drug development and basic science.
Molecular Size and Packaging Capacity
The coding sequence length of a Cas nuclease is a paramount consideration for viral delivery, as it directly dictates compatibility with vector packaging limits. Adeno-Associated Viruses (AAVs), a favored delivery vehicle for in vivo applications, have a stringent payload capacity of approximately 4.7 kilobases (kb) for the entire genetic cassette, including promoters and other regulatory elements [44].
The following table compares the sizes of common CRISPR nucleases, highlighting their compatibility with all-in-one AAV packaging.
| CRISPR Nuclease | Approximate Coding Sequence Length (amino acids) | AAV "All-in-One" Packaging Compatibility | Key Considerations for Delivery |
|---|---|---|---|
| SpCas9 (S. pyogenes) | 1,368 aa [44] [45] | Incompatible (too large) | Requires split intein systems or dual AAVs, increasing complexity [44]. |
| LbCas12a (L. bacterium) | ~1,200-1,300 aa | Incompatible (too large) | Larger than SpCas9; similar delivery challenges [46]. |
| enEbCas12a (E. bacterium) | ~1,100 aa [46] | Compatible (~4.4 kb cassette) | A novel, compact variant enabling all-in-one AAV delivery with a crRNA [46]. |
| Cas12f (CasMINI) | ~400-500 aa [46] | Compatible | Hypercompact size but initially lower efficiency; engineered variants (e.g., exoCasMINI) show improved performance [47]. |
As the data shows, the discovery and engineering of compact Cas variants like enEbCas12a and Cas12f are pivotal advancements, as they bypass the packaging limitations that have historically hampered the use of wild-type Cas9 and Cas12a in single-AAV therapies [46] [47].
Performance and Editing Outcomes
Beyond size, the functional performance of Cas9 and Cas12a varies significantly in terms of editing precision, mutation profiles, and suitability for multiplexing. The following table summarizes key differences based on experimental data.
| Editing Characteristic | Cas9 | Cas12a | Experimental Support |
|---|---|---|---|
| DSB Structure | Blunt ends [2] [10] | Staggered cuts with 5' overhangs [2] [4] | In vitro cleavage assays [10]; Amplicon sequencing in tomato protoplasts [4]. |
| Mutation Profile | Predominantly small indels [4] | Higher frequency of larger deletions [4] | High-throughput amplicon sequencing in tomato [4]. |
| Precision with HDR | Lower precision with ssODN templates [13] | Slightly higher precision with ssODN templates [13] | RNP delivery with ssODN in Chlamydomonas; total editing was similar, but Cas12a produced more precise knock-ins [13]. |
| Multiplexing | Requires multiple gRNAs or complex operons [2] | Native ability to process a crRNA array [2] [46] [10] | Demonstration of crRNA arrays in mammalian cells and plants [2] [10]. |
| Off-Target Profile | Well-characterized; potential for off-targets with mismatches [45] | Potentially higher specificity; off-targets often require mismatches distal from PAM [10] [4] | CIRCLE-seq and amplicon sequencing in maize and tomato [10] [4]. |
Key Experimental Protocols and Workflows
Protocol: In Vivo Gene Editing with a Compact Cas12a Variant via AAV
This protocol is adapted from a study demonstrating the use of the novel enEbCas12a for in vivo genome editing in mice [46].
1. Vector Construction:
- Clone the engineered enEbCas12a coding sequence (enabled by a point mutation for relaxed PAM and increased efficiency) under a constitutive promoter (e.g., EFS promoter) [46].
- Clone a U6 promoter-driven crRNA expression cassette targeting the gene of interest (e.g., Pcsk9) into the same vector backbone.
- Critical Quality Control: Confirm the final vector genome size is within AAV packaging limits (~4.4 kb for the enEbCas12a system) [46].
2. AAV Production and Purification:
- Package the recombinant vector genome into AAV9 capsids (which have broad tropism) using a standard triple-transfection method in HEK293T cells.
- Purify the viral particles via ultracentrifugation and titrate using qPCR to determine genome copies per mL (gc/mL) [46].
3. In Vivo Delivery and Analysis:
- Administration: Inject the AAV-enEbCas12a preparation intravenously into mice.
- Efficiency Assessment: After 14-21 days, harvest target tissues (e.g., liver). Extract genomic DNA and assess editing efficiency at the target locus using next-generation sequencing (NGS) of PCR amplicons to quantify indel percentages [46].
- Phenotypic Readout: For Pcsk9 targeting, monitor a reduction in serum cholesterol levels as a functional indicator of successful gene disruption [46].
Protocol: Comparing Cas9 and Cas12a Editing in Plant Protoplasts
This high-throughput method allows for rapid, quantitative comparison of nuclease performance, as employed in tomato research [4].
1. Target Selection and Construct Design:
- Select overlapping genomic target sites where Cas9 (NGG PAM) and Cas12a (TTTV PAM) target sequences intersect.
- Design and clone sgRNAs (for Cas9) and crRNAs (for Cas12a) into appropriate expression vectors. A Golden Gate Assembly-based system is recommended for efficient crRNA cloning [4].
2. Protoplast Transfection and Editing:
- Isolate protoplasts from the desired plant tissue (e.g., tomato leaf mesophyll).
- Transfect the protoplasts with CRISPR-Cas constructs, using a ribonucleoprotein (RNP) complex or plasmid DNA delivery. A 96-well format can be used for high-throughput screening [4].
- Incubate the transfected protoplasts for 48-72 hours to allow for genome editing.
3. Mutation Analysis and Specificity Assessment:
- On-target Efficiency: Harvest protoplast genomic DNA. Amplify the on-target regions by PCR and subject the amplicons to high-throughput sequencing. Align sequences to the reference genome to calculate indel frequencies and characterize the spectrum of mutations (small indels vs. large deletions) [4].
- Off-target Analysis: Use bioinformatics tools (e.g., Cas-OFFinder) to predict potential off-target sites with a high degree of sequence similarity. Amplify and sequence these loci from the pooled DNA to empirically determine off-target activity for each nuclease [4].
The Scientist's Toolkit: Essential Research Reagents
The table below details key reagents and their applications for overcoming delivery challenges in CRISPR research.
| Research Reagent | Function and Application |
|---|---|
| AAV Serotypes (e.g., AAV9, AAV8) | Viral capsids with distinct tissue tropisms (e.g., AAV9 for broad tissue targeting, AAV8 for hepatocytes) used for in vivo delivery of compact CRISPR systems [46] [44]. |
| Ionizable Lipid Nanoparticles (LNPs) | Synthetic non-viral delivery vehicles that encapsulate and protect CRISPR cargo (RNP, mRNA), enabling efficient in vivo delivery, particularly to the liver [48] [44]. |
| Ribonucleoprotein (RNP) Complex | Pre-assembled complex of Cas protein and guide RNA. Offers rapid activity, reduced off-target effects, and is the preferred cargo for many non-viral delivery methods [48] [44]. |
| crRNA Array | A single transcript containing multiple crRNAs, processed autonomously by Cas12a. Essential for efficient multiplexed genome editing without complex cloning [2] [46]. |
| Golden Gate Assembly System | A modular, highly efficient DNA cloning method for constructing complex CRISPR vectors, such as those containing multiple gRNAs or crRNA arrays [4]. |
| High-Fidelity Cas Variants | Engineered nucleases (e.g., hfCas12Max, enEbCas12a) with point mutations that increase editing specificity and/or efficiency, reducing off-target effects for therapeutic applications [46] [44] [45]. |
The choice between Cas9 and Cas12a is no longer solely about their intrinsic editing properties but is increasingly dictated by the delivery strategy. For applications where all-in-one AAV delivery is paramount, newly characterized compact Cas12a variants like enEbCas12a provide a compelling solution, merging manageable size with robust editing. In contrast, for ex vivo editing or non-viral LNP delivery where cargo size is less restrictive, the higher on-target efficiency of Cas9 and the multiplexing advantage of Cas12a remain key differentiators. Ultimately, the optimal system is project-specific, hinging on a careful balance of target genomic context, desired editing outcome, and the practical constraints of delivering the machinery into the target cell.
Data-Driven Tool Selection: Validating Performance in Complex Editing Scenarios
The choice between CRISPR-Cas9 and CRISPR-Cas12a for genome engineering in model organisms involves critical trade-offs between targetable genomic space, editing precision, mutational outcomes, and suitability for multiplexed applications. The following table synthesizes key performance metrics from direct comparative studies.
Table 1: Direct Comparison of Cas9 and Cas12a Editing Characteristics
| Feature | CRISPR-Cas9 | CRISPR-Cas12a | Supporting Experimental Data |
|---|---|---|---|
| PAM Requirement | 3'-NGG (SpCas9) [4] | 5'-TTTV (LbCas12a) [7] [4] | |
| Targetable Sites | 32x more in coding sequences; 8x more in promoter regions (in C. reinhardtii) [13] | More limited due to stringent PAM [13] | C. reinhardtii; PAM frequency analysis [13] |
| Total Editing Efficiency | Up to 20-30% (with ssODN); Induced more edits at certain loci (e.g., FKB12) [13] | Achieved similar total editing levels of 20-30% (with ssODN) [13] | C. reinhardtii; RNP delivery with ssODN templates at three genomic loci [13] |
| Precision Editing Efficiency | Slightly lower precision in ssODN-templated editing [13] | Higher precision in ssODN-templated editing [13] | C. reinhardtii; RNP delivery with ssODN templates [13] |
| Mutation Profile | Predominantly small insertions/deletions (indels) [4] | Larger deletions [4] | Tomato protoplasts; Amplicon sequencing at 35 target sites [4] |
| Multiplexing Capability | Requires multiple individual gRNAs or complex expression systems [35] | Superior; natively processes its own crRNA arrays from a single transcript [7] [35] | Demonstrated in human cells with Cas12a-derived base editors editing 15 sites simultaneously [35] |
| Off-Target Activity | Can be higher; improved specificity using paired nickases [24] | Generally lower off-target activity reported [4] | Tomato; assessment of 57 predicted off-target sites for LbCas12a [4] |
Detailed Experimental Data and Protocols
The comparative data in Table 1 is derived from rigorous, direct comparisons in various model systems. Below, we detail the key experiments and their methodologies.
Comparative Editing Efficiency and Precision inChlamydomonas reinhardtii
This study provided a head-to-head comparison of Cas9 and Cas12a in the model alga, focusing on editing with single-stranded oligodeoxynucleotide (ssODN) donor templates [13].
- Experimental System: Chlamydomonas reinhardtii [13].
- Key Reagents: Cas9 and Cas12a Ribonucleoproteins (RNPs), ssODN repair templates [13].
- Methodology:
- Target Selection: Overlapping regions at three independent genomic loci were targeted.
- Delivery: Cas9 or Cas12a RNPs were co-delivered with ssODN repair templates into the cells.
- Analysis: The total editing efficiency (percentage of viably recovered cells with edits) and the precision editing efficiency (successful incorporation of the ssODN template) were quantified.
- Outcome: While both systems achieved similarly high total editing levels (20-30%), Cas12a demonstrated a statistically significant, albeit slight, advantage in precision editing [13].
Mutation Pattern Analysis in Tomato Protoplasts
This research directly compared the nature of mutations induced by LbCas12a and SpCas9 in a plant model system [4].
- Experimental System: Tomato protoplasts [4].
- Key Reagents: SpCas9 and LbCas12a expression vectors, respective guide RNA/crRNA expression constructs [4].
- Methodology:
- Targeting: 35 overlapping target sites within the bHLH transcription factor gene family were selected.
- Transfection: Protoplasts were transfected with CRISPR constructs.
- Sequencing: High-throughput amplicon sequencing was performed on the target sites to characterize the spectrum of induced mutations [4].
- Outcome: LbCas12a induced a higher frequency of larger deletions compared to SpCas9, which predominantly created small indels. This characteristic can be advantageous for applications requiring complete gene knockouts [4].
Advanced Engineering and Multiplexing Capabilities
Beyond wild-type nucleases, engineered variants and derived editors highlight the evolving capabilities of each system.
- PAM Expansion for Cas12a: The limited targeting range of wild-type Cas12a (∼1% of a typical genome) has been addressed through directed evolution. The engineered Flex-Cas12a variant recognizes a 5'-NYHV-3' PAM, expanding potential target sites to ∼25% of the human genome while retaining recognition of the canonical TTTV PAM [7].
- Multiplexed Base Editing with Cas12a: Cas12a's innate ability to process a single long transcript into multiple crRNAs makes it ideal for multiplexing. A Cas12a-derived base editor was used to simultaneously edit 15 target sites in human cell lines, a three-fold increase over the state-of-the-art for mammalian cells at the time. This demonstrates a clear advantage for studying polygenic traits or performing genome-scale engineering [35].
The following diagram illustrates the core functional differences in the DNA targeting and cleavage mechanisms between Cas9 and Cas12a, which underpin their performance characteristics.
The Scientist's Toolkit: Essential Reagents and Materials
Table 2: Key Reagents for Comparative CRISPR-Cas Studies
| Reagent / Solution | Function / Description | Example Use in Context |
|---|---|---|
| Ribonucleoproteins (RNPs) | Pre-complexed Cas protein and guide RNA; reduces off-target effects and enables editing in hard-to-transfect cells [13]. | Direct delivery into C. reinhardtii for efficiency comparison [13]. |
| ssODN (single-stranded oligodeoxynucleotide) | Short, single-stranded DNA template for precise homology-directed repair (HDR) [13]. | Co-delivered with RNPs to assess precision editing in C. reinhardtii [13]. |
| Golden Gate Assembly System | A modular, restriction-ligation based cloning method for efficiently assembling multiple DNA fragments [4]. | Used to construct vectors for expressing Cas12a/crRNAs and complex multiplexed gRNA arrays in tomato [4]. |
| LNP-SNAs (Lipid Nanoparticle Spherical Nucleic Acids) | Advanced nanostructure for delivery; enhances cellular uptake, editing efficiency, and reduces toxicity compared to standard LNPs [49]. | A emerging delivery vehicle shown to triple gene-editing success rates in human cell cultures [49]. |
| Prime Editor (PE) Systems | Fuses a Cas9 nickase (H840A) to a reverse transcriptase; enables precise base conversions, insertions, and deletions without double-strand breaks [50]. | PE systems (e.g., PE2, PE3, PE4) represent an alternative pathway for high-precision editing beyond Cas9/Cas12a nucleases [50]. |
| Cas12a-derived Base Editors (e.g., BEACON, enAsBE) | Fuses catalytically dead Cas12a (dCas12a) to a deaminase enzyme; allows single-nucleotide conversions without double-strand breaks [35]. | Enabled multiplexed base editing at 15 sites in human cells, leveraging Cas12a's native crRNA processing [35]. |
The advent of clustered regularly interspaced short palindromic repeats (CRISPR)-CRISPR-associated (Cas) systems has revolutionized genetic engineering, with Cas9 and Cas12a emerging as two of the most prominent tools for genome editing. While both nucleases create double-strand breaks in DNA, they differ significantly in their structure, protospacer adjacent motif (PAM) requirements, and cleavage mechanisms, leading to important differences in their editing specificities. Cas9 requires a G-rich PAM (5'-NGG-3') and is guided by a single guide RNA (sgRNA), utilizing its HNH and RuvC nuclease domains to create a blunt-ended double-strand break. In contrast, Cas12a recognizes a T-rich PAM (5'-TTTV-3') and is guided by a shorter CRISPR RNA (crRNA), using a single RuvC domain to generate staggered ends with 5' overhangs [51].
A critical consideration for therapeutic applications of CRISPR systems is their specificity—the ability to cleave only at intended on-target sites while avoiding unintended off-target modifications. Cas12a exhibits intrinsically higher specificity than Cas9 due to its stricter requirement for guide-target complementarity, particularly in the PAM-distal region [52] [53]. This inherent difference in specificity profiles necessitates robust methods for identifying potential off-target sites across the genome. Among the various techniques developed for this purpose, CIRCLE-seq has emerged as a highly sensitive in vitro method for comprehensive off-target identification, offering advantages over both cell-based methods and other biochemical approaches [51] [54].
This review provides a systematic comparison of CIRCLE-seq and genomic validation studies for assessing the on-target and off-target activities of CRISPR-Cas9 and Cas12a systems, with particular emphasis on their applications in multiplex genome editing research.
CIRCLE-seq Methodology and Workflow
Principles and Technical Implementation
CIRCLE-seq represents a significant advancement in in vitro methods for identifying genome-wide off-target cleavage sites of CRISPR-Cas nucleases. This technique dramatically reduces the background noise associated with previous methods like Digenome-seq by employing a circularized genomic DNA substrate. The fundamental principle involves converting linear genomic DNA into circular molecules, which are then cleaved in vitro by Cas nuclease-guide RNA complexes. Only nuclease-cleaved fragments contain ends suitable for adapter ligation and subsequent sequencing, thereby enriching for potential off-target sites [54].
The key advantage of CIRCLE-seq lies in its exceptional signal-to-noise ratio, which is approximately 180,000-fold better than Digenome-seq. This enhanced sensitivity allows CIRCLE-seq to identify rare off-target sites that might be missed by other methods, while requiring substantially fewer sequencing reads (approximately 4-5 million reads compared to 400-500 million for Digenome-seq) [54]. Furthermore, CIRCLE-seq does not require reference genome sequences, making it applicable to organisms with incomplete genomic data or outbred populations with significant sequence heterogeneity [54].
Table 1: Key Improvements of CIRCLE-seq Over Previous Methods
| Method | Sequencing Reads Required | Background Noise | Sensitivity | Reference Genome Needed |
|---|---|---|---|---|
| CIRCLE-seq | ~4-5 million | Very low | High | No |
| Digenome-seq | ~400-500 million | High | Moderate | Yes |
| GUIDE-seq | Varies | Low (cellular context) | Moderate-high | Yes |
| SITE-seq | ~0.62-2.46 million | Low | Moderate | Yes |
Experimental Workflow
The CIRCLE-seq protocol involves multiple critical steps that contribute to its high sensitivity and specificity for off-target detection.
Diagram 1: CIRCLE-seq Experimental Workflow. The process begins with genomic DNA extraction, followed by shearing, circularization, in vitro cleavage with Cas nuclease, and sequencing of cleaved fragments.
The workflow begins with high-quality genomic DNA extraction from the target cell type or organism. This DNA is mechanically sheared into fragments of optimal size (typically 300-500 bp) to facilitate efficient circularization. The linear fragments are then circularized through intramolecular ligation, creating covalently closed DNA circles that are resistant to background cleavage events. These circularized DNA molecules are incubated with preassembled Cas nuclease-guide RNA ribonucleoprotein (RNP) complexes under optimized reaction conditions. At this stage, the Cas RNP complexes cleave both on-target and off-target sites present in the circular DNA library, linearizing the corresponding circles [54].
Following nuclease treatment, the reaction is stopped, and the DNA is purified. The cleaved (linearized) fragments are selectively enriched while circular DNA is excluded. Sequencing adapters are ligated to the ends of the linearized fragments, which are subsequently amplified and prepared for high-throughput sequencing. The resulting sequencing data is analyzed using specialized bioinformatic pipelines to map cleavage sites across the genome, with cluster-based algorithms identifying sites with significant enrichment of read start positions corresponding to the characteristic cleavage pattern of the Cas nuclease being tested [54].
Comparative Analysis of Cas9 and Cas12a Editing Profiles
Efficiency and Specificity Profiles
Direct comparisons of Cas9 and Cas12a editing efficiencies and specificities reveal distinct profiles that inform their appropriate applications in genome engineering. Studies across multiple systems consistently demonstrate that Cas9 generally achieves higher on-target editing efficiency than Cas12a, while Cas12a exhibits superior specificity with fewer off-target effects.
In a comprehensive study in maize, Cas9-edited plants showed mutation rates of 90-100% in T0 plants, with 63-77% being homozygous or biallelic mutants. In contrast, Cas12a-edited plants showed significantly lower mutation rates of 0-60% in T0 plants, with no homozygous mutants detected [10]. This efficiency advantage of Cas9 was consistent across multiple target sites. However, when assessing specificity through CIRCLE-seq analysis, Cas9 gRNAs showed substantially more potential off-target sites (18 and 67 for two different gRNAs) compared to Cas12a crRNAs (10 and 4 potential off-target sites with ≤4 mismatches) [10].
Table 2: Performance Comparison of Cas9 and Cas12a in Genome Editing
| Parameter | Cas9 | Cas12a | Experimental Context |
|---|---|---|---|
| On-target efficiency | 90-100% mutation rate | 0-60% mutation rate | Maize T0 plants [10] |
| Homozygous/biallelic mutants | 63-77% | 0% | Maize T0 plants [10] |
| Potential off-target sites | 18-67 sites | 10-4 sites (≤4 mismatches) | CIRCLE-seq in maize [10] |
| Precision editing with ssODN | Slightly lower | Higher precision | C. reinhardtii [13] |
| Multiplex editing capability | Requires multiple gRNAs | Native multiplexing with single crRNA array | Various systems [24] |
Notably, when co-delivered with single-stranded oligodeoxynucleotide (ssODN) repair templates, Cas9 and Cas12a ribonucleoproteins (RNPs) induced similar total editing levels (20-30%) in Chlamydomonas reinhardtii, but Cas12a showed slightly higher precision in templated genome editing [13]. This suggests that while Cas9 may be more efficient for gene disruption, Cas12a might offer advantages for precise editing applications.
Molecular Basis for Specificity Differences
The differences in specificity between Cas9 and Cas12a stem from their distinct molecular mechanisms of target recognition and cleavage. Cas9 requires a 20-nucleotide guide-target complementarity and a 5'-NGG PAM, but exhibits tolerance to mismatches, particularly in the PAM-distal region. This tolerance varies depending on the position, type, and number of mismatches, with stretches of complementary nucleotides (especially in the seed region near the PAM) often sufficient to promote cleavage at off-target sites [51].
In contrast, Cas12a demonstrates stricter reliance on guide-target complementarity throughout the target sequence, with particularly low tolerance for mismatches in the PAM-distal region. This characteristic contributes to its higher intrinsic specificity. Additionally, Cas12a's requirement for a 5'-TTTV PAM expands the range of targetable sequences beyond Cas9's 5'-NGG requirement, while simultaneously constraining potential off-target sites to those with this specific PAM configuration [52] [53].
The structural basis for these differences lies in the mechanism of DNA unwinding and R-loop formation. Cas9 undergoes more substantial conformational changes upon PAM recognition, which may contribute to its greater tolerance to mismatches. Cas12a, however, employs a different DNA recognition mechanism that maintains stricter surveillance of guide-target complementarity throughout the binding and cleavage process [51].
Advances in Cas12a Engineering for Enhanced Performance
Protein Engineering Strategies
To address the limitations in Cas12a editing efficiency, several protein engineering approaches have been developed. Structure-guided mutagenesis has yielded hyper-efficient Cas12a variants with significantly improved activity. One notable example is hyperCas12a, which incorporates four mutations (D156R/D235R/E292R/D350R) that enhance DNA binding affinity through increased electrostatic interactions between the positively charged arginine residues and negatively charged DNA backbone [8].
This engineered variant demonstrates substantially improved performance across multiple applications. In gene activation assays, hyperCas12a showed remarkable enhancement, particularly under limiting crRNA conditions where it outperformed wildtype Cas12a by over 300-fold [8]. The variant also exhibited broadened PAM compatibility, recognizing not only canonical TTTA, TTTC, and TTTG PAMs but also non-canonical PAMs including TTTT, CTTA, TTCA, and TTCC [8]. These improvements extended to gene repression and direct genome editing applications, making hyperCas12a a versatile tool for multiplexed genome regulation.
Other engineering efforts have focused on different aspects of Cas12a function. The enAsCas12a variant was developed with mutations that enhance both activity and PAM range, while high-fidelity variants (AsCas12a-HF) incorporate mutations that reduce non-specific DNA contacts, further improving specificity at the potential cost of reduced activity [51].
Guide RNA Engineering Approaches
Parallel to protein engineering, innovations in guide RNA design have significantly boosted Cas12a performance. Recent advances include the development of circular guide RNAs (cgRNAs), which demonstrate dramatically improved stability compared to linear guides. These cgRNAs are engineered using Twister ribozymes that flank the guide sequence, enabling autocatalytic cleavage and subsequent circularization by endogenous RNA ligase RtcB [52].
The covalently closed structure of cgRNAs confers exceptional resistance to exonuclease degradation, resulting in significantly extended half-life in cells. In functional assays, cgRNAs increased the efficiency of Cas12a-based transcription activators and genomic DNA cleavage by approximately 2.1- to 40.2-fold for single gene editing and 1.7- to 2.1-fold for multiplexed gene editing compared to their linear counterparts [52]. This enhancement persisted across multiple cell lines and in vivo mouse liver models, demonstrating the broad utility of this approach.
Another innovative guide RNA engineering strategy incorporates synthetic nucleotides to enhance binding affinity. The incorporation of 2-aminoadenine (base Z) into crRNA replaces the standard Watson-Crick A:T base pairing with a three-hydrogen-bond Z:T pairing, increasing thermal stability and sequence specificity [53]. This Z-crRNA approach significantly improved on-target editing efficiency across multiple genomic loci while maintaining Cas12a's low off-target profile, creating a system that combines Cas9-like efficiency with Cas12a's superior specificity [53].
Table 3: Engineering Strategies to Enhance Cas12a Performance
| Engineering Approach | Key Features | Performance Improvement | References |
|---|---|---|---|
| hyperCas12a | Four mutations (D156R/D235R/E292R/D350R) that enhance DNA binding | >300-fold improvement in activation under low crRNA; broadened PAM recognition | [8] |
| circular gRNAs (cgRNAs) | Covalently closed structure resistant to nucleases | 2.1-40.2-fold increase for single editing; 1.7-2.1-fold for multiplex editing | [52] |
| Z-crRNA | 2-aminoadenine substitution enables 3 H-bonds with thymine | Increased editing from 17% to 65% at tested sites; maintained high specificity | [53] |
| enAsCas12a | Engineered for enhanced activity and expanded PAM recognition | Improved editing efficiency while maintaining specificity | [51] |
Validation Methods and Therapeutic Considerations
Orthogonal Validation of Off-Target Sites
Identification of potential off-target sites via CIRCLE-seq requires validation in relevant cellular models to assess their biological significance. While CIRCLE-seq demonstrates high sensitivity in detecting in vitro cleavage sites, not all biochemical off-targets result in mutations in cellular contexts due to chromatin accessibility, DNA repair mechanisms, and cellular environment factors.
Orthogonal validation methods include targeted deep sequencing, which remains the gold standard for confirming the presence of nuclease-induced indels at putative off-target sites [51]. This approach involves designing primers flanking potential off-target sites identified by CIRCLE-seq and performing high-coverage sequencing to detect mutations in nuclease-treated cells. Other validation methods include GUIDE-seq, which captures off-target breaks through integration of double-stranded oligodeoxynucleotides, and DISCOVER-seq, which exploits DNA repair proteins to mark recently cleaved sites [51].
Studies comparing CIRCLE-seq with cell-based methods have shown strong correlation while highlighting their complementary strengths. CIRCLE-seq identified all off-target sites found by GUIDE-seq for four out of six gRNAs tested, while also detecting numerous additional sites not identified by the cell-based method [54]. Similarly, CIRCLE-seq detected 50 of 53 off-target sites (94%) previously identified by HTGTS, another sensitive cell-based method [54]. These findings suggest that CIRCLE-seq provides comprehensive coverage of potential off-target sites, while cell-based methods identify those most relevant in specific cellular contexts.
Implications for Therapeutic Development
The distinct efficiency and specificity profiles of Cas9 and Cas12a, combined with comprehensive off-target assessment methods like CIRCLE-seq, have significant implications for therapeutic development. For applications requiring high efficiency and where complete characterization of off-target risk is possible, such as ex vivo cell therapies, Cas9 may be preferable. Its well-characterized behavior, high efficiency, and availability of high-fidelity variants make it suitable for these applications [51].
For in vivo therapeutic applications where controlling off-target effects is paramount, Cas12a's intrinsic high specificity offers significant advantages. This is particularly relevant for therapies targeting essential genes or those with narrow therapeutic windows. The emergence of engineered hyper-efficient Cas12a variants further narrows the efficiency gap while maintaining superior specificity [8] [53].
The multiplexing capability of Cas12a is especially valuable for complex genetic engineering applications. Its native ability to process multiple crRNAs from a single transcript simplifies simultaneous targeting of multiple genomic loci, enabling sophisticated genome engineering strategies. This capability has been demonstrated in diverse contexts, including reprogramming of retinal progenitor cells in postnatal mice through simultaneous activation of endogenous Oct4, Sox2, and Klf4 genes [8].
The Scientist's Toolkit: Essential Research Reagents
Table 4: Essential Reagents for CIRCLE-seq and CRISPR Specificity Profiling
| Reagent/Resource | Function | Application Notes |
|---|---|---|
| CIRCLE-seq kit | Comprehensive workflow for off-target identification | Available from commercial suppliers; includes circularization enzymes, controls, and analysis software |
| High-fidelity DNA ligase | Efficient circularization of sheared genomic DNA | Critical for minimizing background; T4 DNA ligase commonly used |
| Cas nuclease (Cas9/Cas12a) | Target DNA cleavage | Recombinantly purified, active protein essential for in vitro cleavage |
| Guide RNA | Target specification | Chemically synthesized or in vitro transcribed; requires purity and proper folding |
| Next-generation sequencing platform | High-throughput sequencing of cleaved fragments | Illumina platforms most commonly used; requires ~4-5 million reads per sample |
| Bioinformatic analysis pipeline | Identification of cleavage sites from sequencing data | Custom scripts or commercial software; CIRCLE-seq analysis tools available |
| Target validation reagents | Orthogonal confirmation of off-target sites | Includes primers for targeted deep sequencing, T7EI assay components, or tracking reagents |
CIRCLE-seq has established itself as a powerful method for comprehensive profiling of CRISPR-Cas nuclease specificities, offering superior sensitivity and practicality compared to earlier techniques. The method's ability to identify potential off-target sites without requiring reference genomes or cellular systems makes it particularly valuable for preclinical assessment of gene editing therapeutics.
The comparative analysis of Cas9 and Cas12a reveals a classic trade-off between efficiency and specificity, with Cas9 generally offering higher on-target activity while Cas12a demonstrates superior specificity and inherent multiplexing capability. Recent engineering advances, including hyper-efficient Cas12a variants, circular guide RNAs, and modified crRNAs, are progressively bridging the efficiency gap while maintaining Cas12a's favorable specificity profile.
For researchers pursuing multiplex genome editing, Cas12a represents an increasingly attractive platform, particularly as engineering improvements address its historical limitations. The combination of comprehensive off-target assessment using CIRCLE-seq, orthogonal validation in relevant models, and selection of appropriately engineered nuclease variants provides a robust framework for developing safe and effective genome editing applications across basic research and therapeutic contexts.
The field of genome editing has been revolutionized by CRISPR technologies, with Cas9 and Cas12a emerging as two of the most powerful tools for precise genetic manipulation. While Cas9 has been the workhorse for numerous breakthroughs, the unique capabilities of Cas12a—particularly for multiplexed gene editing—are now being unlocked through advanced model systems. The recent development of Cas12a-knock-in mice represents a significant technological advancement, providing researchers with a versatile platform for conducting complex genetic screens, disease modeling, and immune cell engineering directly in vivo. This guide objectively compares the performance of these emerging Cas12a murine systems against established Cas9 alternatives, providing researchers with experimental data and methodologies to inform their study designs.
Fundamental Differences: Cas9 vs. Cas12a Systems
Understanding the core molecular distinctions between Cas9 and Cas12a is essential for selecting the appropriate tool for multiplexed screening applications. The table below summarizes key biochemical differences that translate to distinct experimental advantages.
Table 1: Fundamental Characteristics of Cas9 and Cas12a Nucleases
| Feature | Cas9 | Cas12a |
|---|---|---|
| Class & Type | Class 2, Type II [40] | Class 2, Type V [40] |
| PAM Sequence | 5'-NGG-3' (SpCas9) [40] [42] | 5'-TTTV-3' or 5'-TTTN-3' (Ultra variant) [2] [40] |
| Guide RNA | Two-part (crRNA + tracrRNA) or single-guide RNA (sgRNA) [40] | Single crRNA (42-44 nt) [40] |
| DNA Cleavage | Blunt-ended double-strand breaks [42] | Staggered cuts with 5' overhangs [2] [42] |
| Domains | RuvC-like and HNH [40] | RuvC-like only [40] |
| Multiplexing | Requires multiple individual sgRNAs | Intrinsic RNase processes crRNA arrays [17] [55] |
These fundamental differences confer specific experimental advantages. Cas12a's T-rich PAM preference makes it ideal for targeting AT-rich genomic regions that are challenging for Cas9 [2]. The staggered DNA ends created by Cas12a can encourage homology-directed repair (HDR), potentially leading to more predictable editing outcomes compared to the blunt ends generated by Cas9 [2] [42]. Most importantly for screening, Cas12a's intrinsic RNase activity allows it to process a single RNA transcript containing multiple crRNAs into mature guides, significantly simplifying multiplexed gene targeting [17] [55].
Recent pioneering work has established both conditional and constitutive Cas12a-knock-in mice, primarily using the Rosa26 locus for transgene insertion [17] [55]. These models typically employ C57BL/6 backgrounds and express optimized Cas12a variants such as LbCas12a or high-fidelity enhanced AsCas12a (enAsCas12a) [17] [55].
The genetic design places the Cas12a transgene under control of a CAG promoter, with expression regulated by a LoxP-Stop-LoxP (LSL) cassette for Cre-dependent activation [17]. This enables tissue-specific or constitutive Cas12a expression when crossed with appropriate Cre driver lines. The constructs also incorporate nuclear localization signals (NLS) at both N- and C-termini to ensure proper nuclear targeting, along with fluorescent reporters (eGFP or mCherry) for tracking expression [17] [55].
Critically, studies have demonstrated that constitutive expression of Cas12a in these models does not lead to discernible pathology or significant alterations in immune cell populations, confirming the viability of these mice for long-term in vivo studies [17] [55].
Performance Comparison: Cas12a vs. Cas9 in Experimental Systems
Editing Efficiency and Specificity
Direct comparisons of editing efficiency between Cas9 and Cas12a reveal context-dependent performance. In plant systems, Cas9 has demonstrated high efficiency (90-100% mutation rates in T0 plants), while Cas12a showed more variable performance (0-60% under similar conditions) [10]. However, optimized Cas12a systems in tomato protoplasts achieved editing efficiencies up to 86.5% for single-gene targeting, comparable to Cas9 systems [4] [41].
Table 2: Comparative Editing Performance Across Biological Systems
| System/Application | Cas9 Efficiency | Cas12a Efficiency | Notes |
|---|---|---|---|
| Maize Transformation [10] | 90-100% (T0 plants with indels) | 0-60% (T0 plants with indels) | Efficiency varied by target site |
| Tomato Protoplasts [4] | ~31.7% (single target) | ~86.5% (single target) | Target-dependent variability observed |
| Aspergillus niger [41] | 31.7% (single gRNA) | 86.5% (single gRNA) | Both systems achieved ~100% with multiple gRNAs |
| Murine Primary Fibroblasts [55] | Not reported | ~100% (single and multiplexed) | enAsCas12a-KI model with lentiviral crRNA delivery |
| Multiplexed Editing (4 genes) [55] | Requires complex multi-guide systems | ~100% editing for all four targets | Single crRNA array delivery in enAsCas12a-KI MDFs |
Multiplexed Editing Capabilities
For multiplexed screening applications, Cas12a offers distinct advantages. Research using enAsCas12a-knock-in mouse-derived fibroblasts demonstrated simultaneous editing of four genes (Trp53, Bim, Puma, and Noxa) with nearly 100% efficiency from a single crRNA array [55]. This streamlined multiplexing capability arises from Cas12a's intrinsic RNase activity, which processes precursor crRNA transcripts into mature guides without requiring complex multi-guide constructs [17].
In cancer modeling, Cas12a-knock-in mice enabled efficient induction of salivary gland squamous cell carcinoma and lung adenocarcinoma through AAV-mediated delivery of a single crRNA array simultaneously targeting Trp53, Apc, Pten, and Rb1 [17]. This demonstrates the power of Cas12a systems for complex in vivo disease modeling that would require substantially more resources with Cas9-based approaches.
Experimental Protocols for Cas12a-Knock-in Mouse Applications
In Vivo Gene Editing via Lipid Nanoparticles (LNPs)
The Cas12a-knock-in mouse platform enables efficient in vivo gene editing without viral delivery. One validated protocol involves:
- crRNA Design: Design crRNAs targeting desired genomic loci with appropriate TTTV PAM sequences.
- LNP Formulation: Encapsulate crRNA molecules in optimized lipid nanoparticles.
- Administration: Systemically administer LNPs to constitutive Cas12a-expressing mice via intravenous or intraperitoneal injection.
- Validation: Assess editing efficiency 3-7 days post-injection using next-generation sequencing of target tissues [17].
This approach has successfully achieved functional knockout of the transthyretin (TTR) protein in mouse liver, demonstrating therapeutic potential [17].
Multiplexed crRNA Delivery via Adeno-Associated Viruses (AAVs)
For sustained expression and more complex editing applications:
- crRNA Array Design: Concatenate multiple crRNA units separated by direct repeat sequences into a single expression cassette.
- AAV Packaging: Package the crRNA array into AAV particles (serotypes 8 or 9 often used for broad tropism).
- In Vivo Delivery: Administer AAVs to Cas12a-knock-in mice via appropriate routes (intravenous, intramuscular, or local injection).
- Phenotypic Monitoring: Monitor for disease development or molecular changes over time [17].
This method has proven effective for autochthonous cancer modeling through multiplexed tumor suppressor inactivation [17].
Ex Vivo Immune Cell Engineering
Cas12a-knock-in mice provide a superior platform for immune cell engineering:
- Cell Isolation: Harvest primary immune cells (T cells, B cells, bone-marrow-derived dendritic cells) from Cas12a-knock-in mice.
- Retroviral Transduction: Transduce cells with retroviral vectors encoding crRNA arrays targeting multiple immune regulators.
- Adoptive Transfer: Introduce engineered cells into recipient mice for functional studies [17].
This protocol enables efficient multiplexed gene perturbation in primary immune cells without requiring electroporation or transfection [17].
The Researcher's Toolkit: Essential Reagents for Cas12a-Knock-in Mouse Studies
Table 3: Key Research Reagents for Cas12a-Knock-in Mouse Experiments
| Reagent/Tool | Function | Example Application |
|---|---|---|
| LSL-enAsCas12a mice [17] | Constitutive or inducible Cas12a expression | Foundational model for in vivo editing studies |
| LSL-LbCas12a mice [17] | Alternative Cas12a variant with distinct PAM preferences | Targeting different genomic loci |
| crRNA Expression Vectors [17] [55] | Delivery of single or multiplexed guide RNAs | AAV, lentiviral, or retroviral packaging |
| Lipid Nanoparticles (LNPs) [17] | Non-viral crRNA delivery | Transient in vivo gene editing |
| AAV Serotypes [17] | In vivo viral delivery of crRNA arrays | Tissue-specific multiplexed editing |
| CMV-Cre mice [17] [55] | Activation of conditional Cas12a alleles | Generation of constitutive expression models |
| Direct Repeat Vectors [4] | Assembly of crRNA arrays for multiplexing | Simultaneous targeting of multiple genes |
The development of Cas12a-knock-in mice represents a significant expansion of the CRISPR toolbox, particularly for applications requiring multiplexed gene perturbation. While Cas9 remains highly effective for many single-gene editing applications, Cas12a offers distinct advantages for complex genetic screens, disease modeling, and immune cell engineering.
The choice between systems should be guided by specific experimental needs:
- Choose Cas9 for: Established protocols, maximum single-guide efficiency, and projects requiring NGG PAM sites.
- Choose Cas12a-knock-in mice for: Multiplexed screening applications, targeting AT-rich genomic regions, and studies where simplified guide RNA production provides significant experimental advantages.
As the Cas12a mouse platform continues to be adopted and optimized, it is poised to accelerate the deconvolution of complex gene interactions and advance our understanding of polygenic diseases, ultimately bridging the gap between high-throughput screening and physiological relevance in intact organisms.
The discovery of CRISPR-Cas systems has revolutionized biological research, offering an unprecedented ability to edit and regulate genomes with precision. Among the myriad of CRISPR systems, the type II effector Cas9 and the type V effector Cas12a have emerged as two of the most powerful and widely adopted platforms for genome engineering. While Cas9 has dominated the field since its initial application, Cas12a presents distinct molecular characteristics that make it particularly valuable for specific applications, especially multiplex genome editing. More recently, engineered variants of both nucleases have been developed to overcome inherent limitations, providing researchers with an expanded toolbox for sophisticated genetic manipulations. This guide synthesizes current experimental evidence to provide a structured framework for selecting the optimal CRISPR system based on specific research goals, with a particular focus on multiplex editing strategies essential for studying complex genetic networks and pathways.
Molecular Mechanisms and Key Distinctions
Understanding the fundamental differences in the molecular mechanisms of Cas9 and Cas12a is crucial for making an informed choice between these systems.
Guide RNA Architecture and Processing
- Cas9 requires two RNA components for targeting: a CRISPR RNA (crRNA) that contains the target-specific spacer, and a trans-activating crRNA (tracrRNA) that facilitates processing and complex formation. These are often fused into a single-guide RNA (sgRNA) ~100 nucleotides in length [2] [10].
- Cas12a operates with a single, shorter crRNA (~42-44 nucleotides) and does not require a tracrRNA [2]. This simpler architecture contributes to its advantage in multiplexed applications.
DNA Cleavage Mechanisms and Repair Outcomes
- Cas9 possesses two distinct nuclease domains (HNH and RuvC) that cleave opposite strands of the target DNA, generating blunt-ended double-strand breaks immediately upstream of the protospacer adjacent motif (PAM) [2] [10].
- Cas12a utilizes a single RuvC domain to cleave both DNA strands, resulting in staggered cuts with 5' overhangs of 4-5 nucleotides [2] [4]. These "sticky ends" can be advantageous for precise DNA insertion as they potentially facilitate homology-directed repair (HDR) or microhomology-mediated end joining (MMEJ) [2] [4].
Table 1: Fundamental Characteristics of Cas9 and Cas12a
| Feature | Cas9 | Cas12a |
|---|---|---|
| PAM Sequence | 3'-NGG-5' (G-rich) | 5'-TTTV-3' (T-rich) |
| Guide RNA | Two RNAs (crRNA & tracrRNA) or sgRNA (~100 nt) | Single crRNA (~42-44 nt) |
| Cleavage Type | Blunt ends | Staggered ends (5' overhangs) |
| crRNA Processing | Requires host factors (RNase III) or expressed as sgRNA | Built-in RNase activity processes own crRNA arrays |
| Size | ~1368 amino acids (SpCas9) | ~1300 amino acids (LbCas12a) |
Diagram 1: Molecular mechanisms of Cas9 and Cas12a compared
Quantitative Performance Comparison
Direct comparative studies across multiple organisms provide valuable insights into the performance characteristics of Cas9 versus Cas12a.
Editing Efficiency
Editing efficiency shows significant variation depending on the target organism, specific target site, and delivery method:
- In maize, Cas9 demonstrated markedly higher efficiency in T0 generation plants, with 90-100% of edited plants carrying indels and 63-77% being homozygous or biallelic mutants. In contrast, Cas12a editing rates ranged from 0-60% in T0 plants [10].
- In tomato protoplasts, LbCas12a showed similar overall editing efficiency to SpCas9, though this was highly target-dependent. Cas12a induced more and larger deletions than Cas9, which can be advantageous for specific knockout applications [4].
- A study in Chlamydomonas reinhardtii found that Cas9 and Cas12a ribonucleoproteins (RNPs) co-delivered with single-stranded oligodeoxynucleotide (ssODN) templates induced similar total editing levels (20-30%), though Cas12a showed slightly higher precision editing [13].
Mutation Profiles and Specificity
The pattern of induced mutations differs significantly between the two nucleases:
- Cas9 predominantly produces small insertions or deletions (indels) at the target site [4].
- Cas12a tends to generate larger deletions,
- In maize, the CRISPR/Cas9 system tended to edit within the on-target range, while the CRISPR/Cpf1 system preferred to edit in between targets, especially in later generations (T2) [12].
Table 2: Experimental Performance Comparison Across Organisms
| Organism | Cas9 Efficiency | Cas12a Efficiency | Key Findings | Citation |
|---|---|---|---|---|
| Maize | 90-100% (T0) | 0-60% (T0) | Cas9 more efficient in initial generations; Cas12a generates more diverse mutations in later generations | [10] [12] |
| Tomato | Target-dependent | Similar to Cas9, target-dependent | Cas12a produces more and larger deletions; both systems effective | [4] |
| Chlamydomonas | 20-30% total editing | 20-30% total editing | Similar total editing; Cas12a slightly higher precision with ssODN templates | [13] |
| Human Cells | High efficiency | Variable, target-dependent | Engineered Cas12a variants significantly enhance efficiency | [8] |
Multiplex Genome Editing Applications
Multiplex editing—simultaneously targeting multiple genomic loci—represents one of the most significant advantages of CRISPR systems for studying genetic networks and pathways.
Cas12a's Native Multiplexing Capability
Cas12a possesses a unique built-in RNase activity that enables it to process a single long transcript into multiple mature crRNAs [2] [56]. This intrinsic feature simplifies multiplexed targeting as researchers need only express a single array of crRNAs separated by direct repeats, rather than multiple individual guides with separate promoters [56].
This capability has been successfully demonstrated in various systems:
- In mammalian cells, tandem expression of Cas12a and a crRNA array from a single Pol II promoter enabled cleavage of five target genes and transcriptional regulation of ten additional targets concurrently [56].
- In plants, Cas12a's multiplexing capability has been harnessed for targeting multiple genes simultaneously, though efficiency can vary [4] [12].
Cas9 Multiplexing Strategies
While Cas9 lacks intrinsic crRNA processing capability, several synthetic biology approaches have been developed to enable multiplexing:
- tRNA-based systems: gRNAs are flanked by tRNA sequences that are processed by endogenous cellular machinery [56] [12].
- Ribozyme-based systems: Self-cleaving ribozymes (Hammerhead and HDV) flank each gRNA, enabling precise excision from a long transcript [56].
- Csy4-based processing: The bacterial RNase Csy4 is co-expressed to cleave at specific recognition sites inserted between gRNAs [56].
Diagram 2: Decision framework for selecting CRISPR systems in multiplex editing
Engineered Variants: Overcoming Natural Limitations
Protein engineering has addressed several limitations of wild-type Cas nucleases, resulting in variants with enhanced capabilities.
Enhanced Cas12a Variants
- hyperCas12a: Created through structure-guided engineering of four residues (D156R/D235R/E292R/D350R), this LbCas12a variant shows significantly enhanced efficacy for gene activation, particularly at low crRNA concentrations. It maintains crRNA processing capability while demonstrating minimal off-target effects compared to the wild-type system [8].
- Flex-Cas12a: This variant incorporates six mutations (G146R/R182V/D535G/S551F/D665N/E795Q) that relax PAM requirements from TTTV to NYHV (Y = C/T; H = A/C/T), expanding potential target sites from ~1% to over 25% of the genome while maintaining robust nuclease activity [7].
Cas9 Variants
While not the focus of this guide, it's worth noting that numerous engineered Cas9 variants with altered PAM specificities, enhanced fidelity, and other improved characteristics have been developed, providing additional options for targeting challenging genomic regions.
Table 3: Engineered Cas12a Variants and Their Applications
| Variant | Key Mutations | PAM Preference | Advantages | Best Applications |
|---|---|---|---|---|
| hyperCas12a | D156R/D235R/ E292R/D350R | TTTV (canonical) with improved non-canonical recognition | Enhanced efficiency, especially at low crRNA levels; improved activation & repression | Multiplexed editing in challenging systems; in vivo applications |
| Flex-Cas12a | G146R/R182V/ D535G/S551F/ D665N/E795Q | NYHV (expanded) | Dramatically expanded targeting range (~25% of genome) | Targeting previously inaccessible loci; therapeutic applications |
| enAsCas12a | E174R/S542R/ K548R | Expanded PAM range | Earlier engineered variant with broader PAM recognition | General use with expanded targeting |
Experimental Protocols for Comparative Assessment
Protocol 1: Side-by-Side Comparison in Plant Systems
This protocol is adapted from studies in maize and tomato that directly compared Cas9 and Cas12a efficiency [4] [10].
Vector Construction:
- For Cas9: Clone maize codon-optimized Cas9 under a ZmUbi promoter, with gRNAs driven by RNA Pol III promoters (U6 or U3).
- For Cas12a: Clone maize codon-optimized LbCas12a under ZmUbi promoter, with crRNAs expressed as an array under Pol III promoter or as a single transcript with ribozyme flanking.
- Include a selectable marker (e.g., Bar gene for bialaphos resistance) for plant selection.
Transformation and Analysis:
- Perform Agrobacterium-mediated transformation of immature embryos (maize) or protoplast transfection (tomato).
- Regenerate plants under selection pressure.
- Genotype T0 plants and subsequent generations by sequencing target loci.
- Quantify editing efficiency as percentage of plants with indels and characterize mutation patterns.
Protocol 2: Assessing Multiplex Editing Efficiency in Mammalian Cells
This protocol evaluates the performance of hyperCas12a for multiplexed gene activation [8].
Cell Line Engineering:
- Generate a HEK293T reporter cell line with genomically integrated TRE3G-GFP.
- Co-transfect plasmids encoding hyperdCas12a-miniVPR fusion and TRE3G promoter-targeting crRNA array.
Efficiency Assessment:
- Analyze GFP activation via flow cytometry 72 hours post-transfection.
- Compare performance to wild-type dCas12a by gating for low BFP population (proxy for low crRNA conditions).
- For endogenous targets, measure mRNA levels of target genes via RT-qPCR.
The Scientist's Toolkit: Essential Research Reagents
Table 4: Key Reagent Solutions for CRISPR Multiplex Editing Research
| Reagent/Category | Specific Examples | Function & Application Notes |
|---|---|---|
| Expression Vectors | pCAMBIA3301-based (plants), pYPQ210 (maize), CL7/Im7 tagged (purification) | Delivery of Cas genes and guide RNAs; selection marker expression |
| Cas Variants | SpCas9, LbCas12a, hyperCas12a, Flex-Cas12a | Endonuclease effector proteins with varying PAM requirements and efficiencies |
| Guide RNA Systems | tRNA-gRNA arrays (Cas9), direct repeat crRNA arrays (Cas12a), ribozyme-flanked guides | Enable multiplexed targeting; choice depends on Cas protein and organism |
| Promoter Systems | ZmUbi (maize), 35S (plants), U6/U3 (Pol III), CAG (mammalian) | Drive expression of Cas proteins and guide RNAs; tissue-specificity considerations |
| Delivery Methods | Agrobacterium-mediated (plants), protoplast transfection, viral delivery (mammalian) | Introduction of CRISPR components into target cells or organisms |
| Analysis Tools | CIRCLE-seq, CasOFFinder, CRISPOR, amplicon sequencing | Design guides and assess on-target efficiency and off-target effects |
The choice between Cas9, Cas12a, and their engineered variants should be guided by specific research requirements:
Choose Cas9 when: Maximum editing efficiency in initial generations is critical (particularly in plants), when targeting G-rich genomic regions, or when working with well-established protocols in your model organism.
Choose Cas12a when: Native multiplexed editing is a priority, when targeting AT-rich genomic regions, when staggered ends are desirable for HDR, or when vector simplicity and size are limiting factors.
Choose engineered variants (hyperCas12a/Flex-Cas12a) when: Targeting efficiency with wild-type nucleases is insufficient, when targeting loci with restricted PAM availability, or when working in challenging systems with low guide RNA expression.
As CRISPR technologies continue to evolve, the optimal choice will increasingly depend on the specific genomic targets, delivery constraints, and desired editing outcomes of each experiment. Researchers are encouraged to conduct pilot comparisons when embarking on new projects, as performance can vary significantly across target sites and biological systems.
Conclusion
The choice between CRISPR-Cas9 and Cas12a for multiplex editing is not a matter of superiority but of strategic application. Cas9 currently offers higher efficiency in many contexts and a more mature toolkit, while Cas12a provides a more streamlined path for complex, multi-gene edits due to its intrinsic crRNA processing and potential for higher precision. The ongoing development of hyper-efficient engineered Cas12a variants is rapidly closing the efficiency gap. The future of multiplex editing lies in leveraging the strengths of each system—using Cas9 for high-efficiency single or dual edits and Cas12a for sophisticated multi-gene circuits and in vivo modeling. This will be accelerated by improved delivery methods and AI-driven design, paving the way for advanced cell therapies and the deconvolution of complex disease mechanisms in biomedical research.
References
- 1. Cas9 versus Cas12a/Cpf1: Structure-function comparisons ... [https://pubmed.ncbi.nlm.nih.gov/29790280/]
- 2. Cas9 vs Cas12a: A Revolutionary Chapter in Genome ... [https://invivobiosystems.com/crispr/cas9-vs-cas12a/]
- 3. CRISPR-Cas Technology as a Revolutionary Genome Editing ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10707817/]
- 4. Comparison of Cas12a and Cas9-mediated mutagenesis ... [https://www.nature.com/articles/s41598-024-55088-4]
- 5. Key thermodynamic characteristics of Cas9 and Cas12a ... [https://www.sciencedirect.com/science/article/abs/pii/S0006291X25006060]
- 6. High-performance CRISPR-Cas12a genome editing for ... [https://www.nature.com/articles/s41467-020-17209-1]
- 7. Directed evolution expands CRISPR–Cas12a genome- ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12266133/]
- 8. Multiplexed genome regulation in vivo with hyper-efficient ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9035114/]
- 9. How to Choose the Right Cas Variant for Every CRISPR ... [https://www.synthego.com/guide/how-to-use-crispr/cas-nuclease-variants/]
- 10. Activities and specificities of CRISPR/Cas9 and Cas12a ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6320322/]
- 11. PAM-adjacent DNA flexibility tunes CRISPR-Cas12a off- ... [https://www.nature.com/articles/s41598-025-87565-9]
- 12. Comparative Study between the CRISPR/Cpf1 (Cas12a ... [https://www.mdpi.com/2077-0472/11/5/429]
- 13. Comparison of CRISPR/Cas9 and Cas12a for gene editing ... [https://www.sciencedirect.com/science/article/pii/S2211926424004089]
- 14. CRISPR-Cas12a: Functional overview and applications - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC7090318/]
- 15. Cas12a - Wikipedia [https://en.wikipedia.org/wiki/Cas12a]
- 16. A CRISPR Cas12a/Cpf1 strategy to facilitate robust multiplex ... [https://fungalbiolbiotech.biomedcentral.com/articles/10.1186/s40694-025-00196-7]
- 17. Cas12a-knock-in mice for multiplexed genome editing ... [https://www.nature.com/articles/s41551-025-01371-2]
- 18. Comparison of CRISPR-Cas9/Cas12a Ribonucleoprotein ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6973557/]
- 19. CRISPR-Cas12a induced DNA double-strand breaks are ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9684475/]
- 20. Development of a CRISPR/Cas12a genome editing ... [https://www.sciencedirect.com/science/article/pii/S2405805X25001565]
- 21. Design of highly functional genome editors by modelling ... [https://www.nature.com/articles/s41586-025-09298-z]
- 22. CRISPR Clinical Trials: A 2025 Update [https://innovativegenomics.org/news/crispr-clinical-trials-2025/]
- 23. CRISPR-Cas9/Cas12a systems for efficient genome ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11521959/]
- 24. Applications of multiplexed CRISPR–Cas for genome ... [https://www.nature.com/articles/s12276-025-01500-6]
- 25. Massively parallel knock-in engineering of human T cells - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC11260498/]
- 26. Cas12a-knock-in mice for multiplexed genome editing ... [https://pubmed.ncbi.nlm.nih.gov/40114032/]
- 27. Increasing Gene Editing Efficiency via CRISPR/Cas9 [https://www.mdpi.com/2227-9059/12/1/119]
- 28. CRISPR 101: Cas9 vs. The Other Cas(s) [https://blog.addgene.org/crispr-101-cas9-vs.-the-other-cass]
- 29. CRISPR/Cas systems: Delivery and application in gene therapy [https://pmc.ncbi.nlm.nih.gov/articles/PMC9723151/]
- 30. Programmable Multiplex Genome Editing: Innovations in ... [https://www.sciepublish.com/article/pii/680]
- 31. AAV-mediated direct in vivo CRISPR screen identifies ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5614841/]
- 32. CRISPR screening by AAV episome-sequencing (CrAAVe- ... [https://www.nature.com/articles/s41593-025-02043-9]
- 33. CRISPR Clinical Trials to Follow [https://www.synthego.com/blog/crispr-clinical-trials/]
- 34. CRISPR-Cas9/Cas12a Systems for efficient genome ... [https://www.pnnl.gov/publications/crispr-cas9cas12a-systems-efficient-genome-editing-and-large-genomic-fragment]
- 35. Precision multiplexed base editing in human cells using ... [https://www.nature.com/articles/s41467-025-59653-x]
- 36. Increasing Cas12a- and Cas9-mediated deletion sizes with ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12581816/]
- 37. A multiplex CRISPR/Cas9 platform for fast and efficient ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5512712/]
- 38. The Complete Guide to Understanding CRISPR sgRNA [https://www.synthego.com/guide/how-to-use-crispr/sgrna/]
- 39. CRISPR Guide [https://www.addgene.org/guides/crispr/]
- 40. Alt-R CRISPR-Cas9 vs Cas12a systems [https://eu.idtdna.com/pages/technology/crispr/crispr-genome-editing/Alt-R-systems]
- 41. CRISPR-Cas9/Cas12a systems for efficient genome ... [https://www.frontiersin.org/journals/bioengineering-and-biotechnology/articles/10.3389/fbioe.2024.1452496/full]
- 42. What's the Difference Between Cas9 and Cas12a Nucleases? [https://www.the-scientist.com/what-s-the-difference-between-cas9-and-cas12a-nucleases-72481]
- 43. Off-target effects in CRISPR/Cas9 gene editing - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10034092/]
- 44. CRISPR Delivery Methods: Cargo, Vehicles, and Challenges [https://www.synthego.com/blog/delivery-crispr-cas9/]
- 45. A Visual Guide to Genome Editors [https://press.asimov.com/articles/crispr-guide]
- 46. When size matters: A novel compact Cas12a variant for in vivo ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11253977/]
- 47. CMN Weekly (25 July 2025) - Your Weekly CRISPR ... [https://crisprmedicinenews.com/news/cmn-weekly-25-july-2025-your-weekly-crispr-medicine-news/]
- 48. Challenges in delivery systems for CRISPR-based genome ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8316527/]
- 49. Scientists just made CRISPR three times more effective [https://www.sciencedaily.com/releases/2025/09/250907024543.htm]
- 50. Emerging trends in prime editing for precision genome ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12322275/]
- 51. Assessing and advancing the safety of CRISPR-Cas tools [https://www.nature.com/articles/s41467-023-35886-6]
- 52. Engineered circular guide RNAs boost CRISPR/Cas12a [https://pmc.ncbi.nlm.nih.gov/articles/PMC10288759/]
- 53. Harnessing noncanonical crRNA for highly efficient ... [https://www.nature.com/articles/s41467-024-48012-x]
- 54. CIRCLE-seq: a highly sensitive in vitro screen for genome- ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5924695/]
- 55. Advancing the genetic engineering toolbox by combining ... [https://www.nature.com/articles/s41467-025-56282-2]
- 56. Multiplexed CRISPR technologies for gene editing and ... [https://www.nature.com/articles/s41467-020-15053-x]