Prime Editing vs. Base Editing: A Precision Genome Engineering Showdown
This article provides a comprehensive comparison of two leading precision genome editing technologies: base editing and prime editing.
Prime Editing vs. Base Editing: A Precision Genome Engineering Showdown
Abstract
This article provides a comprehensive comparison of two leading precision genome editing technologies: base editing and prime editing. Tailored for researchers, scientists, and drug development professionals, it explores the foundational mechanisms of each tool, their methodological applications across research and therapy, persistent challenges with current optimization strategies, and rigorous validation approaches. By synthesizing the latest advancements, including next-generation editors with minimized errors, this analysis serves as a critical resource for selecting the appropriate editing strategy for specific experimental or therapeutic goals, from correcting point mutations to installing complex genetic variants.
Understanding the Core Mechanisms of Precision Editing
Base editing is a precise genome editing technology that enables the direct, irreversible conversion of one DNA base pair to another without making double-stranded DNA breaks (DSBs) [1] [2]. This technology represents a significant advancement over traditional CRISPR-Cas9 methods, as it avoids the error-prone repair pathways that often lead to unintended insertions or deletions (indels) [3] [4].
The core principle of base editing involves the chemical modification of a target nucleobase within the genome. By fusing a catalytically impaired CRISPR-Cas protein to a nucleobase-modifying enzyme, base editors can directly change the chemical structure of a single DNA base, leading to a permanent and precise point mutation [1] [5].
Core Components and Mechanisms
A functional DNA base editor consists of three primary molecular components that work in concert [4]:
- A Catalytically Impaired Cas Protein: Typically, a "dead" Cas9 (dCas9, which cannot cut DNA) or a Cas9 "nickase" (nCas9, which cuts only one DNA strand) is used. This protein is responsible for recognizing and binding to the target DNA sequence, guided by a gRNA [1] [4].
- A Nucleobase-Modifying Enzyme: This enzyme performs the chemical conversion on the target base. The specific enzyme determines the type of base edit [1].
- A Guide RNA (gRNA): This RNA molecule directs the Cas protein to the specific genomic locus intended for editing [6] [4].
Upon binding to the target DNA, the Cas protein locally unwinds the DNA duplex, creating a single-stranded DNA "bubble" known as an R-loop [1] [5]. This exposes a short stretch of single-stranded DNA, making it accessible to the tethered deaminase enzyme. The editing activity is confined to a narrow "editing window," typically about 5 nucleotides wide, within this displaced strand [1] [6].
Cytosine Base Editors (CBEs)
Cytosine Base Editors catalyze the conversion of a C•G base pair to a T•A base pair [1] [5]. The first base editor, BE1, was developed by fusing dCas9 to a rat APOBEC1 cytidine deaminase enzyme [1] [4]. This enzyme deaminates cytosine (C) in single-stranded DNA to uracil (U). In the cell, uracil is read as thymine (T) during DNA replication or repair. Subsequent generations of CBEs have incorporated key improvements [4]:
- BE2: Fusion of the uracil DNA glycosylase inhibitor (UGI) protein to block base excision repair, which would otherwise reverse the U•G intermediate back to C•G [1] [5].
- BE3: Use of a Cas9 nickase to cut the non-edited DNA strand, encouraging the cell to use the U-containing strand as a template during repair, thereby increasing editing efficiency [1] [4].
- BE4: Incorporation of a second UGI copy and optimized linkers to further reduce unwanted byproducts like C-to-G or C-to-A conversions and minimize indel formation [4].
Diagram of the CBE editing mechanism for converting C•G to T•A.
Adenine Base Editors (ABEs)
Adenine Base Editors perform the reverse conversion, changing an A•T base pair to a G•C base pair [4] [5]. The development of ABEs was a significant engineering challenge, as no natural DNA adenine deaminases were known. Researchers used directed evolution to create a DNA-capable adenine deaminase, called TadA, from a naturally occurring RNA adenine deaminase (TadA) [4] [5]. This engineered enzyme deaminates adenine (A) in DNA to inosine (I). The cellular machinery then interprets inosine as guanine (G), leading to an A•T to G•C conversion after repair and replication [5]. ABEs also utilize a Cas9 nickase to improve editing efficiency [4].
Diagram of the ABE editing mechanism for converting A•T to G•C.
Quantitative Comparison of Base Editing Systems
The following table summarizes the key characteristics and performance metrics of major base editor systems, highlighting the evolution of their design and efficiency.
Table 1: Evolution and performance of major base editor systems
| Base Editor | Key Components | Base Conversion | Average Editing Efficiency | Indel Frequency | Notable Features |
|---|---|---|---|---|---|
| BE1 [1] | dCas9-APOBEC1 | C•G → T•A | 0.8–7.7% (in cells) | N/A | First-generation CBE; low efficiency in cells |
| BE3 [1] [4] | nCas9-APOBEC1-UGI | C•G → T•A | ~37% (average across 6 loci) | ~1.1% | Nickase activity and UGI boost efficiency |
| BE4/BE4max [4] | nCas9-APOBEC1-2xUGI (with Gam & codon optimization) | C•G → T•A | 4.2–6.0x improvement over BE3 | 1.5–2.0x reduction vs BE3 | Higher product purity; reduced indels & byproducts |
| Target-AID [1] | nCas9-CDA1-UGI | C•G → T•A | Comparable to BE3 | Not specified | Uses PmCDA1 deaminase; slightly shifted editing window |
| ABE7.10 [4] | nCas9-TadA(v7.10) | A•T → G•C | ~53% | ~1.2% | First-generation ABE; highly specific A-to-G conversion |
| ABEmax [4] | nCas9-TadA(v7.10) with improved NLS & codons | A•T → G•C | Significantly improved over ABE7.10 | Comparable to ABE7.10 | Optimized for mammalian cell expression & nuclear delivery |
| ABE8e [6] [4] | nCas9-TadA-8e (V106W) | A•T → G•C | ~590x faster deamination than TadA in ABE7.10 | No significant increase | Greatly improved editing speed & efficiency at difficult sites |
Experimental Protocols for Base Editing
A standard protocol for evaluating base editing efficiency involves the following key steps [6]:
- gRNA Design and Library Construction: Design gRNAs targeting the genomic loci of interest. For high-throughput analysis, a lentiviral library of gRNA-target pair constructs can be generated. The target sequence typically includes the 20-nucleotide protospacer, the PAM sequence, and flanking genomic sequences.
- Cell Transduction and Editor Expression:
- Culture human cell lines (e.g., HEK293T) engineered to stably express the base editor (e.g., ABE7.10 or BE4).
- Transduce cells with the lentiviral gRNA library at a low multiplicity of infection (MOI ~0.3) to ensure most cells receive a single gRNA. Maintain cells under selective pressure (e.g., puromycin) and induce base editor expression (e.g., with doxycycline) for a defined period, typically around 8 days.
- Harvesting and Sequencing:
- Harvest genomic DNA from the transduced cell population.
- Perform deep amplicon sequencing of the target sites using PCR primers specific to the surrogate target sites. Aim for high sequencing coverage (e.g., >1000 reads per gRNA) to ensure robust data.
- Data Analysis:
- Process sequencing reads to quantify base editing efficiency and outcomes for each gRNA.
- Calculate the percentage of reads containing the desired base conversion (C-to-T or A-to-G) at each position within the editing window (typically positions 3-10 in the protospacer).
- Analyze the frequency of other outcomes, such as bystander edits (unintended edits within the window), transversions (e.g., C-to-G), and indels.
Table 2: Key research reagents and solutions for base editing experiments
| Reagent/Solution | Function in Experiment | Example & Notes |
|---|---|---|
| Base Editor Plasmid | Provides the genetic code for the base editor protein. | Plasmids for BE4max, ABE8e, etc.; often codon-optimized for human cells [4]. |
| gRNA Expression Vector | Drives the expression of the guide RNA that targets the editor to the DNA site. | Can be cloned into lentiviral backbones for delivery [6]. |
| Lentiviral Packaging System | Produces viral particles to deliver editor and gRNA constructs into cells. | Essential for hard-to-transfect cells and in vivo delivery [7]. |
| Cell Line | Provides the cellular environment for editing. | Commonly used: HEK293T, HeLa, or patient-derived iPSCs [6]. |
| Delivery Agent | Facilitates introduction of editors into cells. | Lipofectamine (for plasmids), electroporation (for RNPs), or AAV/LNPs (for in vivo) [7]. |
| Deep Sequencing Service | Quantifies editing efficiency and outcomes at target sites. | Critical for accurate measurement of base conversion rates and byproducts [6]. |
| UNG Inhibitor (UGI) | Critical component of CBEs to prevent repair of U•G intermediates. | Expressed as part of the CBE fusion protein [1] [5]. |
Comparative Analysis with Prime Editing
While base editing excels at making precise transition mutations (purine-to-purine or pyrimidine-to-pyrimidine swaps), its scope is limited to four of the twelve possible base-to-base conversions and it cannot install insertions or deletions cleanly [3] [8]. Prime editing was developed to overcome these limitations.
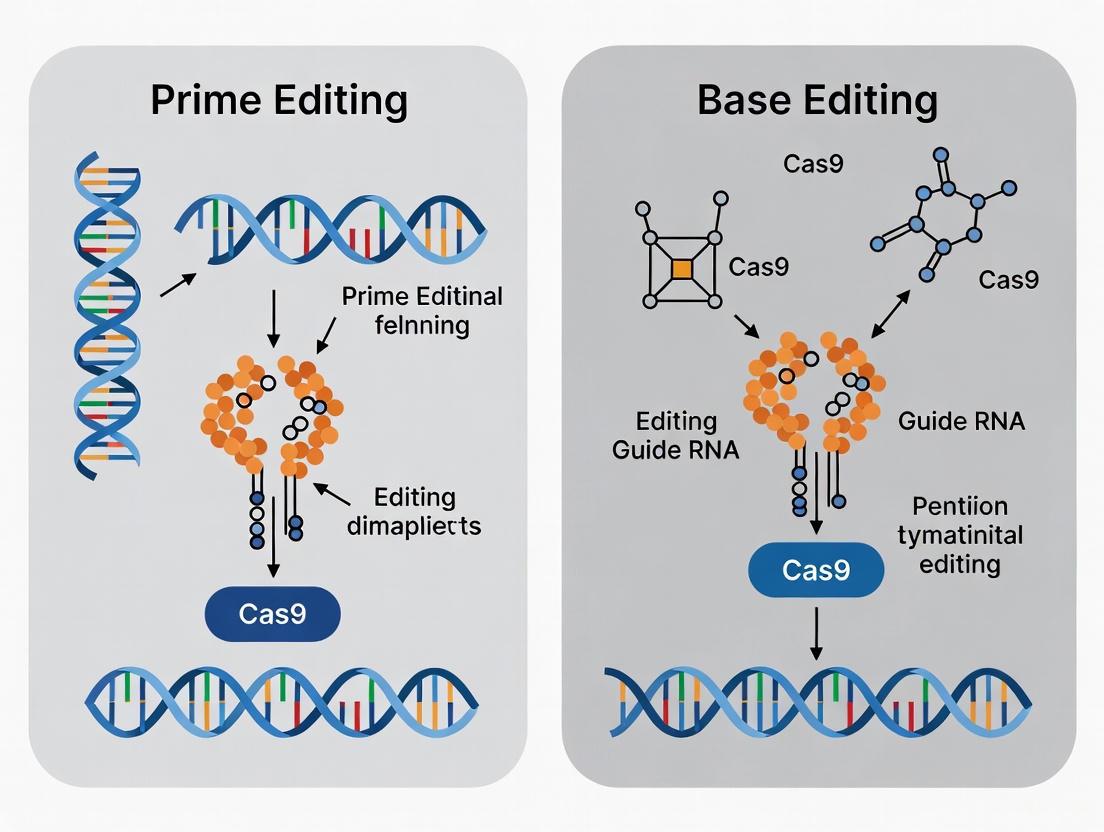
Prime editing uses a catalytically impaired Cas9 nickase fused to a reverse transcriptase (RT), programmed with a prime editing guide RNA (pegRNA) [8] [2]. The pegRNA both specifies the target site and contains a template for the new DNA sequence. The system nicks one DNA strand and uses the RT to "copy" the edited sequence from the pegRNA template into the genome. This "search-and-replace" capability allows prime editors to make all 12 possible base-to-base conversions, as well as small insertions and deletions, without requiring DSBs [8].
Table 3: Key distinctions between base editing and prime editing
| Feature | Base Editing | Prime Editing |
|---|---|---|
| Core Machinery | dCas9/nCas9 + Deaminase + (UGI for CBE) | nCas9 + Reverse Transcriptase |
| Guiding Molecule | standard gRNA | prime editing gRNA (pegRNA) |
| DNA Cleavage | Single-strand nick (or none) | Single-strand nick |
| Editing Scope | C•G → T•A and A•T → G•C (transitions only) | All 12 base-to-base changes, plus insertions and deletions |
| Theoretical Targetability | Limited to specific transition mutations within a editing window. | Could correct up to 89% of known pathogenic human genetic variants [3]. |
| Primary Byproducts | Bystander edits within the activity window; some indels. | Potentially lower bystander edits; can still generate indels [8]. |
| Major Challenge | Restricted scope of edits; bystander mutations. | Large size complicating delivery; variable efficiency [3] [8]. |
Base editing establishes a powerful paradigm for precision genome editing through direct chemical conversion of DNA bases. Its ability to efficiently install point mutations without inducing double-strand breaks makes it a transformative technology for both basic research and therapeutic development, with several base-edited therapies already in clinical trials [3] [7]. However, the limited scope of edits it can make and the challenge of bystander mutations are key considerations for its application.
The choice between base editing and prime editing is context-dependent. Base editing often achieves higher efficiencies for the specific transition mutations it catalyzes. In contrast, prime editing offers vastly greater versatility, capable of correcting a wider range of genetic defects, albeit sometimes with lower efficiency and more complex vector delivery requirements [8]. Together, these technologies provide researchers and clinicians with an expanding toolkit for precise genetic modification, each with distinct advantages suited to different experimental or therapeutic goals.
The advent of CRISPR-Cas systems revolutionized genetic engineering, but the initial reliance on double-strand breaks (DSBs) introduced challenges for precision editing, including unpredictable insertions, deletions, and chromosomal rearrangements [9]. Base editing emerged as a breakthrough technology that enables precise, irreversible single-base conversions without creating DSBs, offering significant advantages for both basic research and therapeutic development [10] [7]. This architecture-focused guide examines the evolution of base editing platforms from foundational Cytosine Base Editors (CBEs) and Adenine Base Editors (ABEs) to the more recent Glycosylase Base Editors (GBEs), providing researchers with a structured comparison of their mechanisms, performance characteristics, and optimal applications within the broader context of precision genome editing.
Architectural Blueprints of Base Editing Systems
Core Architecture and Mechanism
Base editors are sophisticated protein machines that combine the targeting capability of CRISPR systems with the chemical activity of cellular enzymes to directly rewrite single DNA nucleotides. Their core architecture consists of several essential components working in concert.
Programmable DNA-Targeting Module: All base editors utilize a catalytically impaired Cas9 nickase (nCas9) containing a D10A mutation that inactivates the RuvC domain, enabling only single-strand nicking rather than double-strand breaks [10] [9]. This nCas9 is guided to specific genomic loci by a single-guide RNA (sgRNA) and requires a protospacer adjacent motif (PAM) sequence for target recognition.
Deaminase Engine: The heart of the base editing system is a nucleotide deaminase enzyme that catalyzes the chemical conversion of one base to another. This enzyme acts on single-stranded DNA exposed during the R-loop formation when Cas9 binds its target [10].
Accessory Domains: Depending on the editor type, additional protein domains are included to modulate DNA repair pathways and favor desired editing outcomes. These include Uracil Glycosylase Inhibitor (UGI) in CBEs to prevent uracil excision, or uracil-DNA glycosylase (UNG) in GBEs to promote base excision repair [10].
The following diagram illustrates the fundamental mechanism shared by base editors, highlighting how the R-loop formation enables deaminase activity on single-stranded DNA.
Architectural Evolution: From CBEs to GBEs
The development of base editors represents a continuous refinement of protein architecture to expand editing capabilities and improve outcomes.
First-Generation Editors: CBEs and ABEs The original CBE architecture (BE3, BE4) featured a fusion of nCas9 (D10A) with a cytidine deaminase (typically APOBEC1) and one or two copies of UGI [11]. Simultaneously, Target-AID systems employed activation-induced cytidine deaminase (AID) instead of APOBEC1, demonstrating similar functionality with variations in editing window preferences [11]. ABEs represented a significant engineering achievement, utilizing a laboratory-evolved Escherichia coli transfer RNA adenosine deaminase (ecTadA) that was redesigned through directed evolution to deaminate adenine in DNA rather than its natural RNA substrate [10].
Advanced Editors: GBEs GBEs emerged from the observation that removing UGI from BE3 increased C-to-A and C-to-G substitutions [10]. By strategically replacing UGI with uracil-DNA glycosylase (UNG), GBE architecture promotes base excision repair that leads to transversion mutations rather than transitions [10]. Recent studies assessing multiple GBE platforms in rice, tomato, and poplar revealed that each GBE has distinct sequence preferences and editing efficiencies across plant species, indicating specialized rather than universal application [10].
Comparative Analysis of Base Editor Architectures
Structural Components and Editing Outcomes
Table 1: Architectural Components and Base Conversion Capabilities
| Editor Type | Core Components | Deamination Activity | Primary Base Conversion | Secondary Conversions | DNA Repair Manipulation |
|---|---|---|---|---|---|
| CBE | nCas9 (D10A) + Cytidine Deaminase (APOBEC1/AID) + UGI | Cytosine → Uracil | C•G to T•A | Minimal C to G/A (<5%) [10] | UGI inhibits UNG-mediated base excision repair |
| ABE | nCas9 (D10A) + engineered tRNA adenosine deaminase (TadA) | Adenine → Inosine | A•T to G•C | Extremely low indel formation [10] | Cellular repair naturally favors inosine incorporation |
| GBE | nCas9 (D10A) + Cytidine Deaminase + UNG (replaces UGI) | Cytosine → Uracil → Abasic site | C•G to G•C or C•G to A•T | Significant indels possible [10] | UNG promotes base excision repair, enabling transversions |
Performance Metrics and Experimental Data
Table 2: Experimental Performance Characteristics in Mammalian and Plant Systems
| Parameter | CBE | ABE | GBE |
|---|---|---|---|
| Editing Window | ~5 nucleotides, ~15 bp upstream of PAM [11] | ~5 nucleotides, ~15 bp upstream of PAM [10] | ~5 nucleotides, position-dependent [10] |
| Typical Efficiency | 55% in mouse Dmd gene (BE3) [11] | High efficiency with minimal indels [10] | Variable efficiency across species [10] |
| Editing Purity | Moderate (bystander editing common) [10] | High (minimal bystander editing) [10] | Low to moderate (indel formation) [10] |
| Sequence Preference | APOBEC1: TC context; AID: WRC (W=A/T, R=A/G) [11] | Minimal sequence context preference [10] | Platform-dependent preferences [10] |
| Multiplexing Capacity | Demonstrated (3 auditory genes in mice) [11] | Demonstrated in various systems | Limited data available |
| Reported Indel Rates | 1-10% in various studies [11] | <1% [10] | 5-15% [10] |
DNA Repair Pathways Determine Editing Outcomes
The cellular response to base editor activity ultimately determines editing success, with different pathways recruited depending on the editor architecture.
Resolution Pathways for Base Editing Intermediates
The diagram below illustrates how cellular DNA repair mechanisms process the intermediate mismatches created by different base editors, leading to their distinct editing outcomes.
Key Experimental Workflow for Base Editor Evaluation
The following workflow represents a standard methodology for testing and validating base editor performance in cellular systems, as referenced in multiple studies [10] [11].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents and Resources for Base Editing Research
| Reagent Type | Specific Examples | Function/Application | Delivery Methods |
|---|---|---|---|
| Base Editor Plasmids | BE3, BE4, ABE7.10, Target-AID | Express full editor system in cells | Viral vectors, transfection |
| Cas9 Nickase Variants | SpCas9 (D10A), SaCas9 (D10A) | DNA targeting with single-strand nicking | mRNA, RNP complexes |
| Deaminase Enzymes | APOBEC1, AID, evolved TadA | Catalyze base conversion reactions | As fusion proteins with nCas9 |
| Repair Modulator Domains | UGI, UNG | Manipulate DNA repair pathways | As fusion proteins with nCas9 |
| Guide RNA Scaffolds | sgRNA, pegRNA (for prime editing) | Target specificity and edit encoding | In vitro transcription, synthetic |
| Delivery Vehicles | AAV, Lentivirus, LNPs | Efficient cellular delivery | Zygote injection, somatic delivery |
| Analysis Tools | NGS platforms, TIDE, T7E1 | Edit characterization and quantification | Post-editing application |
Base editors represent a remarkable achievement in protein engineering, offering researchers a suite of tools for precision genome manipulation. The architectural progression from CBEs to ABEs and GBEs has progressively expanded the scope of possible edits from transitions to transversions. CBEs remain valuable for C-to-T conversions but struggle with bystander editing in dense cytosine regions. ABEs demonstrate exceptional purity and efficiency for A-to-G conversions with minimal indel formation. GBEs unlock novel transversion editing but with increased complexity in outcome prediction and higher indel rates.
The future of base editing architecture will likely focus on refining editing precision, expanding targetable sequences through engineered Cas variants with altered PAM requirements, and developing systems with minimized off-target effects. As these architectures mature, their integration with emerging technologies like prime editing will provide researchers with an increasingly sophisticated toolkit for precise genetic manipulation, accelerating both basic research and therapeutic development.
In the quest to correct disease-causing genetic mutations, researchers have historically faced a fundamental trade-off: the efficiency of making genetic changes versus the precision of those changes. Traditional CRISPR-Cas9 nucleases create double-strand DNA breaks (DSBs), which efficiently disrupt genes but frequently produce unpredictable insertions/deletions (indels) and other unwanted mutations through error-prone repair processes [12] [9]. Base editing emerged as a breakthrough that avoids DSBs by directly converting one base to another (C-to-T or A-to-G), but its applications are restricted to four transition mutations and it often creates unintended "bystander" edits to nearby bases [12] [13] [9]. Prime editing represents a transformative "search-and-replace" technology that combines the programmability of CRISPR systems with the precision of reverse transcription, enabling versatile genetic modifications without DSBs or the limitations of base editing [12] [8] [14].
Table: Comparison of Major Genome Editing Technologies
| Technology | Editing Scope | DSB Formation | Key Limitations |
|---|---|---|---|
| CRISPR-Cas9 Nuclease | Gene disruption via indels | Yes | Unpredictable repair outcomes, chromosomal rearrangements [12] [9] |
| Base Editing | C-to-T, G-to-A, A-to-G, T-to-C transitions | No | Restricted to 4 base transitions; bystander edits; cannot make transversions [12] [13] [9] |
| Prime Editing | All 12 base-to-base conversions, insertions, deletions | No | Variable efficiency across sites/cell types; large cargo size [12] [8] [14] |
The Architectural Framework of Prime Editing
Core Components and Mechanism
The prime editing system functions as a highly specialized molecular machine composed of two primary components: (1) the prime editor protein, a fusion of a Cas9 nickase (H840A) and an engineered reverse transcriptase (RT), and (2) the prime editing guide RNA (pegRNA), which both specifies the target genomic locus and encodes the desired edit [12] [8] [13]. Unlike traditional CRISPR guide RNAs, pegRNAs contain additional functional regions: a primer binding site (PBS) that anchors the system to the nicked DNA, and a reverse transcriptase template (RTT) that provides the new genetic information to be written into the genome [13].
The editing mechanism occurs through a sophisticated multi-step biochemical process. First, the Cas9 nickase domain recognizes the target DNA sequence adjacent to a protospacer adjacent motif (PAM) and creates a single-strand nick in the DNA. The exposed 3' end then hybridizes with the PBS region of the pegRNA. Next, the reverse transcriptase utilizes the RTT as a template to synthesize a DNA flap containing the desired edit. Finally, cellular repair machinery resolves this intermediate structure, incorporating the edited strand into the genome [12] [8] [13]. For particularly challenging edits, researchers often employ an additional nicking sgRNA (in PE3 systems) to create a nick in the non-edited strand, thereby encouraging the cell to use the edited strand as a repair template and increasing editing efficiency [12] [8].
Evolution of Prime Editing Systems
Since the initial development of PE1, prime editors have undergone significant optimization through protein engineering and mechanistic refinements. The transition from PE1 to PE2 involved engineering the reverse transcriptase component with five mutations (D200N/T306K/W313F/T330P/L603P) that enhanced its thermostability, processivity, and affinity for RNA-DNA hybrid substrates [12] [14]. PE3 systems further improved efficiency by incorporating an additional sgRNA that nicks the non-edited strand to bias cellular repair toward the edited sequence [12] [8]. More recent versions (PE4-PE7) have integrated additional enhancements including engineered pegRNAs with stabilizing RNA motifs, suppression of mismatch repair pathways, and compact reverse transcriptases optimized through phage-assisted continuous evolution (PACE) [12] [14]. The latest systems, including PEmax and PE6 variants, achieve markedly higher editing efficiencies (up to 80-95% in some cell types) while reducing off-target effects [12] [14].
Table: Evolution of Prime Editor Systems
| Editor Version | Key Innovations | Typical Editing Efficiency | Primary Applications |
|---|---|---|---|
| PE1 | Foundational Cas9 nickase-RT fusion | ~10-20% | Proof-of-concept demonstrations [12] |
| PE2 | Engineered RT with 5 stability mutations | ~20-40% | Broad research applications [12] |
| PE3/3b | Additional nicking sgRNA | ~30-50% | Challenging genomic contexts [12] [8] |
| PE4/5 | MMR inhibition (MLH1dn) | ~50-80% | Therapeutic development [12] |
| PE6 variants | Evolved compact RTs, epegRNAs | ~70-90% | In vivo applications, large inserts [12] [14] |
Comparative Performance Analysis: Prime Editing vs. Base Editing
Precision and Specificity Metrics
Direct comparative studies reveal distinct performance characteristics between prime editing and base editing technologies. In a study targeting the cystic fibrosis-causing mutation R785X, adenine base editing (ABE) achieved higher editing efficiency but prime editing demonstrated superior precision by avoiding bystander edits [14]. Base editors frequently exhibit off-target activity due to the deaminase enzymes (APOBEC1, TadA) acting on non-targeted cytosines or adenines within a 4-5 nucleotide editing window, with bystander mutation rates sometimes exceeding 50% at certain loci [12] [9]. In contrast, prime editing requires three independent hybridization events (spacer binding, PBS annealing, and flap resolution), creating a higher barrier to off-target editing and resulting in significantly cleaner editing profiles [9].
Recent advances in protein engineering have substantially improved prime editing precision. MIT researchers developed engineered Cas9 variants that destabilize the original DNA strand after cutting, encouraging the cell to incorporate the corrected sequence and reducing error rates from approximately 1 in 7 edits to as low as 1 in 543 edits in high-precision mode [15]. This enhanced vPE system demonstrates that strategic protein engineering can address earlier limitations in prime editing fidelity without adding operational complexity [15].
Editing Scope and Versatility
The most significant distinction between these technologies lies in their editing capabilities. Base editors are fundamentally limited to four transition mutations (C→T, G→A, A→G, T→C) and cannot address transversions (C→A, C→G, etc.), insertions, or deletions [12] [9]. Prime editing's modular architecture enables all 12 possible base-to-base conversions, plus targeted insertions (routinely up to 40+ bp), deletions, and combinations thereof [12] [8] [13]. This versatility makes prime editing particularly valuable for researching and potentially treating diverse genetic disorders, as different conditions require distinct types of genetic corrections.
Analysis of pathogenic mutations in the ClinVar database indicates that approximately 16,000 small deletions could potentially be repaired using prime editing for therapeutic purposes, vastly expanding the addressable disease landscape compared to base editing [14]. Furthermore, specialized prime editing systems like twinPE can facilitate large DNA insertions by installing recombinase "landing pads," enabling kilobase-scale genetic modifications without double-strand breaks - a capability entirely beyond the scope of base editing technologies [14].
Table: Quantitative Performance Comparison in Model Systems
| Performance Metric | Base Editing | Prime Editing |
|---|---|---|
| Editing Scope | 4 transition mutations | All 12 point mutations, insertions, deletions [12] [13] |
| Bystander Edits | Common in editing window (4-5 bases) | Rare [12] [9] |
| DSB Formation | Very low | Very low [12] [9] |
| Max Efficiency in HEK293T | Up to 70% (varies by site) | Up to 95% with latest systems [12] [14] |
| Indel Formation | Low (<1-5%) | Low (<1-5%), further reducible with engineered editors [12] [9] |
| Therapeutic Applications | Single nucleotide transitions | Broad range of mutation types [16] [14] |
Experimental Workflows and Methodologies
Prime Editing Workflow for Precise Genome Modification
Implementing prime editing requires careful experimental design and optimization across multiple parameters. The process begins with pegRNA design, where researchers must select appropriate spacer sequences (typically ~20 nt) targeting the genomic locus of interest, design the reverse transcriptase template (RTT, 25-40 nt) encoding the desired edit, and include a primer binding site (PBS, 10-15 nt) with sufficient complementarity to initiate reverse transcription [13]. Contemporary approaches often incorporate engineered pegRNAs (epegRNAs) with stabilizing RNA secondary structures (evopreQ, mpknot, or G-quadruplex motifs) at their 3' termini to protect against exonucleolytic degradation and improve editing efficiency by 3-4 fold [8] [9].
The next critical phase involves delivery system selection based on the target cell type. For mammalian cell lines, plasmid-based transfection remains common, with systems like PEmax offering improved editing efficiency through optimized nuclear localization signals and codon usage [14]. For therapeutically relevant primary cells or in vivo applications, viral delivery (typically dual AAV vectors due to cargo size constraints) or lipid nanoparticles (LNPs) represent the leading approaches [16] [7]. Recent advances include split-intein systems that reconstitute full-length editors post-delivery and vPE systems with enhanced precision profiles [15] [7].
Following delivery, researchers must modulate cellular environment to favor edit persistence, particularly by temporarily inhibiting mismatch repair pathways which can reverse prime edits. The PE4 and PE5 systems address this challenge by incorporating dominant-negative MLH1 (MLH1dn), increasing editing efficiency 2-3 fold by preventing the cellular machinery from rejecting the edited strand [12] [14]. Finally, comprehensive analysis of editing outcomes through next-generation sequencing of both on-target and potential off-target sites is essential to validate editing efficiency and specificity [17] [9].
The Scientist's Toolkit: Essential Research Reagents
Table: Essential Reagents for Prime Editing Research
| Reagent Category | Specific Examples | Function & Notes |
|---|---|---|
| Prime Editor Plasmids | PE2, PEmax, PE6 variants | Engineered Cas9-RT fusions with varying efficiency/size profiles [12] [14] |
| pegRNA Expression Systems | epegRNA backbones, xr-pegRNAs | Protect against degradation; improve efficiency 3-4 fold [8] [9] |
| Delivery Vehicles | LNPs, AAV vectors (dual), electroporation systems | Critical for challenging cell types; consider cargo size limitations [13] [16] |
| MMR Inhibitors | MLH1dn, small molecules | Enhance editing persistence; integrated in PE4/5 systems [12] [14] |
| Analysis Tools | NGS assays, computational prediction algorithms | Validate on-target efficiency, detect off-target effects [17] [9] |
The choice between prime editing and base editing technologies depends fundamentally on the specific research objectives and genetic context. Base editing offers superior efficiency for straightforward transition mutations (C→T, A→G) where bystander edits are not a concern, particularly in contexts like introducing premature stop codons or correcting specific point mutations [9] [7]. Prime editing provides a vastly more versatile platform capable of addressing diverse mutation types (transversions, insertions, deletions) with exceptional precision, making it ideal for modeling complex genetic diseases and developing therapeutic approaches for disorders requiring specific sequence corrections [12] [14].
As both technologies continue evolving—with base editors expanding their targeting scope and reducing bystander effects, and prime editors achieving higher efficiencies and more compact configurations—researchers now possess an increasingly sophisticated toolkit for precision genome engineering. The recent FDA approval of the first prime editing-based Investigational New Drug (PM359 for chronic granulomatous disease) underscores the transition of these technologies from basic research to therapeutic development, heralding a new era of precision genetic medicine [16].
The emergence of prime editing represents a paradigm shift in precision genome engineering, offering an unprecedented level of control over genetic modifications. At the heart of this technology lies the prime editing guide RNA (pegRNA), a multifaceted molecule that serves as both a targeting device and a blueprint for desired edits. Unlike earlier genome editing technologies that rely on double-strand breaks (DSBs)—which can lead to unpredictable insertions, deletions, and chromosomal rearrangements—prime editing enables precise modifications without inducing DSBs or requiring donor DNA templates [8] [12].
This advancement addresses significant limitations of both nuclease-based CRISPR systems and base editing technologies. While base editing circumvented the need for DSBs, it remains restricted to specific base transitions (C-to-T or A-to-G) and can cause unintended bystander edits to adjacent nucleotides [8] [3]. Prime editing, in contrast, theoretically enables all 12 possible base-to-base conversions, along with targeted insertions and deletions, dramatically expanding the scope of editable genetic variants [13]. This capability positions prime editing as a transformative tool for both basic research and therapeutic development, with computational models suggesting it could correct up to 89% of known pathogenic human genetic variants [3].
The following comparison guide examines how the pegRNA-driven prime editing system achieves its precision, directly comparing its performance against base editing technologies through analysis of experimental data and methodological approaches.
Molecular Mechanisms: pegRNA vs. Base Editing Systems
Architecture of the pegRNA
The pegRNA is a sophisticated engineered RNA molecule that fundamentally expands upon the capabilities of traditional single-guide RNAs (sgRNAs) used in CRISPR systems. It consists of four essential components that work in concert to enable precise editing [13]:
- Target Sequence: A ~20-nucleotide spacer that directs the Cas9 nickase to the specific DNA target site through complementary base pairing.
- Scaffold Sequence: Maintains the secondary structure necessary for binding to the Cas9 nickase enzyme.
- Primer Binding Site (PBS): A 10-15 nucleotide sequence that anneals to the nicked DNA strand, serving as an initiation point for reverse transcription.
- Reverse Transcription Template (RTT): Encodes the desired edit along with flanking homology, typically 25-40 nucleotides in length.
The complete pegRNA molecule generally ranges from 120-145 nucleotides, significantly longer than standard sgRNAs, which presents both functional advantages and delivery challenges [13].
Comparative Mechanism of Action
Table 1: Mechanism Comparison Between Prime Editing and Base Editing
| Editing Feature | Prime Editing | Base Editing |
|---|---|---|
| Core Components | Cas9 nickase-reverse transcriptase fusion + pegRNA | Cas9-deaminase fusion + sgRNA |
| DNA Cleavage | Single-strand nick | Single-strand nick or no cut |
| Edit Template | Encoded in pegRNA's RTT | No template required |
| Editing Window | Flexible, determined by RTT length | Restricted (4-5 nucleotides) |
| Primary Editing Outcomes | All 12 base transitions, insertions, deletions | C-to-T or A-to-G conversions only |
| Cellular Repair Pathway | Flap equilibrium and resolution | DNA mismatch repair |
The fundamental difference between these technologies lies in their molecular mechanisms. Base editors utilize deaminase enzymes that directly convert one base to another—cytidine deaminases for C-to-T transitions and adenosine deaminases for A-to-G transitions [8] [3]. These enzymes are fused to Cas9 proteins that often nick the non-edited strand to improve efficiency. However, this approach is constrained by a small editing window of four to five nucleotides and can result in bystander edits where adjacent bases within the window are unintentionally modified [8].
In contrast, the prime editing mechanism is more complex but offers greater precision and versatility [8] [12] [13]:
- The pegRNA directs the prime editor (a Cas9 nickase-reverse transcriptase fusion) to the target DNA site.
- Cas9 nickase creates a single-strand break in the non-target DNA strand.
- The exposed 3' end hybridizes with the PBS sequence of the pegRNA.
- Reverse transcriptase synthesizes DNA using the RTT as a template, incorporating the desired edit.
- Cellular repair mechanisms resolve the resulting DNA flap structure, incorporating the edit into the genome.
- An optional second nick on the non-edited strand (in PE3 systems) can enhance editing efficiency.
This "search-and-replace" capability allows prime editing to install precise mutations without being confined to specific base transitions or a restricted editing window [13].
Figure 1: pegRNA Architecture and Prime Editing Mechanism. The pegRNA contains four critical regions that guide the prime editing complex to perform precise genetic modifications.
Performance Comparison: Quantitative Analysis of Editing Outcomes
Editing Efficiency and Precision
Recent advancements in prime editing systems have substantially improved editing efficiencies, making the technology competitive with base editing approaches while maintaining superior precision. The evolution from initial PE1 systems to optimized versions like PEmax has been instrumental in this progress.
Table 2: Evolution of Prime Editing Systems and Their Performance
| Editor Version | Key Improvements | Editing Efficiency | Notable Features | Reference |
|---|---|---|---|---|
| PE1 | Foundational system | ~10-20% in HEK293T cells | Initial proof-of-concept | [12] |
| PE2 | Engineered reverse transcriptase | ~20-40% in HEK293T cells | Improved stability and processivity | [8] [12] |
| PE3 | Additional sgRNA for nicking non-edited strand | ~30-50% in HEK293T cells | Enhanced efficiency via strand bias | [8] [12] |
| PEmax | Further optimized RT and Cas9 | Up to 80% in multiple cell lines | Codon optimization, nuclear localization | [18] [19] |
| PE6 | Compact RT variants, engineered Cas9 | ~70-90% in HEK293T cells | Improved delivery compatibility | [12] |
Experimental data demonstrates that optimized prime editing systems can achieve remarkable efficiencies under ideal conditions. One systematic optimization approach combining stable genomic integration of prime editors via the piggyBac transposon system with enhanced promoters and lentiviral delivery of engineered pegRNAs (epegRNAs) achieved up to 80% editing efficiency across multiple loci and cell lines [18]. In human pluripotent stem cells (hPSCs)—notoriously challenging for genome editing—this system still achieved substantial editing efficiencies of up to 50% [18].
The most significant efficiency improvements have come from addressing cellular repair mechanisms that often reverse prime edits. Implementation of mismatch repair (MMR) inhibition through dominant-negative MLH1 (MLH1dn) in PE4 and PE5 systems has dramatically increased editing yields by preventing the cellular machinery from recognizing and reversing the edited strand [12] [19]. In benchmarked experiments, MMR-deficient cell lines reached near-perfect editing efficiencies of ~95% for certain edits when using stabilized epegRNAs over extended time periods [19].
Specificity and Off-Target Profiles
While both base editing and prime editing show favorable off-target profiles compared to traditional CRISPR-Cas9 nucleases, they exhibit different specificity challenges:
Base editing limitations [8] [3]:
- Bystander edits: Unintended modifications of adjacent bases within the editing window
- DNA/RNA off-target activity: Deaminase enzymes can exhibit promiscuous activity
- Restricted editing scope: Limited to four of twelve possible base transitions
Prime editing advantages [8] [19]:
- Minimal bystander editing: Precise installation of only the intended changes
- Reduced off-target effects: No significant increase in unwanted mutations observed
- Broad editing scope: Capable of all 12 base-to-base conversions, insertions, and deletions
Quantitative assessments of editing precision in high-throughput screens have demonstrated that prime editing produces highly specific outcomes with minimal errors. One benchmarked study evaluating thousands of epegRNA-target pairs found generally low error frequencies (median errors <4%) alongside high-efficiency precise editing [19].
Experimental Protocols for pegRNA-Based Editing
High-Efficiency Prime Editing Workflow
Achieving robust prime editing outcomes requires careful experimental design and optimization. The following protocol, adapted from highly efficient implementations, provides a framework for researchers seeking to compare prime editing with alternative approaches [18] [19]:
Step 1: pegRNA Design and Selection
- Design pegRNAs with varying PBS lengths (10-15 nt) and RTT lengths (10-30 nt)
- Incorporate evopreQ1 or mpknot motifs at the 3' end to create epegRNAs with enhanced stability
- Utilize computational tools like PEGG (Prime Editing Guide Generator) for high-throughput design
Step 2: Delivery System Optimization
- For in vitro applications: Employ piggyBac transposon system for stable genomic integration of editor components
- Use strong constitutive promoters (CAG, EF1α) for robust editor expression
- Deliver epegRNAs via lentiviral vectors for sustained expression
Step 3: MMR Inhibition
- Co-express dominant-negative MLH1 (MLH1dn) to suppress mismatch repair
- Consider using MMR-deficient cell lines (e.g., MLH1 knockout) for maximum efficiency
Step 4: Extended Expression and Selection
- Maintain edited cells for 2-4 weeks to allow edit accumulation
- Implement fluorescence-based sorting if editors include reporter genes (e.g., mCherry)
Step 5: Validation and Sequencing
- Employ amplicon sequencing with unique molecular identifiers for accurate efficiency quantification
- Assess both on-target efficiency and potential off-target effects through targeted sequencing
Figure 2: High-Efficiency Prime Editing Workflow. The optimized experimental protocol for achieving robust prime editing outcomes incorporates stable editor expression and mismatch repair inhibition.
High-Throughput Screening with Prime Editing Sensors
For comparative studies evaluating multiple genetic variants, prime editing sensor systems enable highly parallel assessment of editing efficiency and functional impact [20] [19]. This approach involves:
Library Design:
- Create pegRNA libraries targeting diverse variants using computational tools like PEGG
- Couple each pegRNA with a synthetic "sensor" site that recapitulates the native target architecture
- Include multiple pegRNAs per variant with different PBS/RTT combinations
Implementation:
- Clone pegRNA-sensor pairs into lentiviral vectors
- Transduce at low multiplicity of infection (MOI ~0.7) to ensure single-copy integration
- Culture cells for extended periods (28 days) with periodic sampling
Analysis:
- Sequence sensor sites to quantify editing efficiency over time
- Correlate pegRNA abundance with phenotypic outcomes in dropout screens
- Compare endogenous editing efficiency with sensor readouts for validation
This sensor-based approach has been successfully applied to study over 1,000 TP53 variants, demonstrating prime editing's capability for functional genomics at scale [20].
Research Reagent Solutions for Prime Editing
Successful implementation of prime editing requires specific reagents and optimization approaches. The following table details essential components and their functions based on current best practices.
Table 3: Essential Research Reagents for Prime Editing Experiments
| Reagent Category | Specific Examples | Function | Considerations |
|---|---|---|---|
| Editor Expression Plasmids | pCMV-PE2, pCMV-PEmax-P2A-hMLH1dn | Express the Cas9 nickase-reverse transcriptase fusion | PEmax shows improved efficiency over PE2; include MMR inhibition |
| pegRNA Delivery Vectors | Lentiviral vectors with U6 promoter | Enable sustained pegRNA expression | Ensure proper folding; avoid excessive secondary structure |
| Stability-Enhanced pegRNAs | epegRNAs with tevopreQ1, mpknot motifs | Protect against 3' degradation | Improve efficiency 3-4-fold over standard pegRNAs |
| Delivery Systems | piggyBac transposon, Lentivirus, AAV, LNPs | Facilitate cellular uptake of editing components | piggyBac ideal for stable integration; dual AAV for large editors |
| MMR Inhibitors | MLH1dn (dominant-negative) | Block mismatch repair to enhance editing | Can increase efficiency 2-3 fold; essential for certain edits |
| Validation Tools | Amplicon sequencing with UMIs | Accurately quantify editing efficiency | Essential for detecting precise edits and byproducts |
The pegRNA represents a fundamental advancement in precision genome editing, serving as an all-in-one targeting and template mechanism that enables precise genetic modifications without double-strand breaks. While base editing remains a valuable tool for specific base transitions, prime editing powered by optimized pegRNAs offers superior versatility, capable of installing all 12 base substitutions along with insertions and deletions.
Current experimental data demonstrates that highly optimized prime editing systems can achieve editing efficiencies of 50-80% in standard cell lines and up to 95% in mismatch repair-deficient systems [18] [19]. These efficiencies, combined with minimal off-target effects and a broad editing scope, position pegRNA-driven prime editing as the preferred technology for precise genetic modifications in research and therapeutic development.
As delivery methods continue to improve and editor designs evolve, the pegRNA blueprint will undoubtedly expand the boundaries of precision genetic medicine, potentially enabling correction of the vast majority of known disease-causing genetic variants.
The advent of CRISPR-Cas9 technology revolutionized genetic engineering by providing researchers with an unprecedented ability to target specific genomic loci. However, the reliance on double-strand breaks (DSBs) introduced significant limitations, including unpredictable repair outcomes, p53-mediated cellular stress, and undesirable insertions/deletions (indels) [8] [12]. These challenges prompted the development of two major DSB-free precision editing platforms: base editing and prime editing. While both technologies represent substantial advances over conventional CRISPR-Cas9 systems, they operate through fundamentally distinct molecular mechanisms with complementary strengths and limitations for research and therapeutic applications [13].
Base editing, pioneered in 2016, utilizes fusion proteins comprising a catalytically impaired Cas protein and a deaminase enzyme to enable direct chemical conversion of one DNA base to another without inducing DSBs [21]. Prime editing, introduced in 2019, employs a more complex mechanism combining a Cas9 nickase with a reverse transcriptase, programmed through specialized guide RNAs to execute precise genetic changes [8] [13]. This article provides a comprehensive mechanistic comparison of these two innovative technologies, examining their operational principles, performance characteristics, and optimal applications within the context of modern genetic research and therapeutic development.
Molecular Mechanisms: A Tale of Two Architectures
Base Editing: Targeted Chemical Conversion
Base editors function through a relatively straightforward mechanism that combines targeted DNA binding with enzymatic base conversion. The core architecture consists of a catalytically impaired Cas protein (typically a nickase) fused to a deaminase enzyme. Cytosine base editors (CBEs) utilize cytidine deaminases to convert cytosine (C) to thymine (T), while adenine base editors (ABEs) employ engineered adenine deaminases to convert adenine (A) to guanine (G) [13] [21]. These editors operate within a defined "editing window" of approximately 4-8 nucleotides in the spacer region, with efficiency influenced by both the specific deaminase variant and the local sequence context [8] [21].
The base editing process initiates when the Cas component binds to the target DNA sequence specified by the guide RNA. Unlike standard CRISPR-Cas9 systems, base editors do not create double-strand breaks. Instead, the deaminase enzyme performs a chemical conversion on a specific base within the editing window, creating an intermediate mismatch. Cellular repair machinery then resolves this mismatch, preferentially using the edited strand as a template, ultimately resulting in a permanent base substitution [21]. This mechanism enables highly efficient conversion of specific base pairs while avoiding the pitfalls associated with DSB repair, though it is constrained to particular transition mutations (C•G to T•A or A•T to G•C) within a narrow editing window [8].
Prime Editing: Search-and-Replace Genomics
Prime editing employs a more versatile but complex mechanism that enables a broader range of precise edits without requiring DSBs or donor DNA templates. The system comprises two key components: a prime editor protein and a prime editing guide RNA (pegRNA). The prime editor is a fusion protein consisting of a Cas9 nickase (H840A variant) connected to an engineered reverse transcriptase (RT) from the Moloney murine leukemia virus [8] [14]. The pegRNA serves a dual function, both directing the complex to the target genomic locus and encoding the desired edit through its 3' extension, which contains a primer binding site (PBS) and a reverse transcriptase template (RTT) [13].
The prime editing mechanism unfolds through a series of coordinated molecular events. First, the Cas9 nickase component binds to the target DNA and creates a single-strand nick, exposing a 3' hydroxyl group on the DNA. This exposed end hybridizes with the PBS region of the pegRNA, forming a primer-template complex that initiates reverse transcription. The RT enzyme then synthesizes DNA using the RTT as a template, generating a 3' flap containing the desired edit. Cellular repair processes subsequently resolve this intermediate structure, incorporating the edited strand into the genome [8] [14]. This sophisticated mechanism supports all 12 possible base-to-base conversions, small insertions, deletions, and combinations thereof, substantially expanding the scope of programmable genome editing beyond what base editors can achieve [13].
Performance Comparison: Efficiency, Precision, and Versatility
Editing Capabilities and Limitations
Table 1: Fundamental Editing Capabilities of Base Editing vs. Prime Editing
| Editing Feature | Base Editing | Prime Editing |
|---|---|---|
| Base Substitutions | C→T, G→A, A→G, T→C (4 conversions) | All 12 possible base-to-base conversions [13] |
| Small Insertions | Not supported | Up to 100+ bp demonstrated [22] |
| Small Deletions | Not supported | Yes, with high precision [8] |
| Editing Window | Narrow (typically 4-8 nucleotides) [21] | Broad, adjustable via pegRNA design [22] |
| Bystander Edits | Common challenge in editing window [8] | Minimal with proper design [23] |
| DSB Formation | Very low | Low (PE3 systems slightly higher) [8] |
| PAM Dependency | Requires canonical NGG for SpCas9 | Requires canonical NGG for SpCas9 [8] |
The data reveal fundamental trade-offs between these technologies. Base editors offer superior efficiency for the specific transitions they catalyze but lack versatility. Prime editors provide remarkable editing flexibility but historically faced efficiency challenges, particularly for certain genomic contexts [23]. Recent advances have substantially improved prime editing efficiency through engineered pegRNAs (epegRNAs), optimized editor architectures (PEmax, PE6 variants), and mismatch repair inhibition [23] [14].
Quantitative Performance Metrics
Table 2: Experimental Performance Comparison Across Applications
| Application Context | Base Editing Efficiency | Prime Editing Efficiency | Key Notes |
|---|---|---|---|
| Point Mutation Correction (HEK293T) | Up to 70% for compatible edits [21] | 20-50% (PE3 systems) [8] | PE efficiency highly dependent on pegRNA design |
| CFTR F508del Correction | Not applicable (requires deletion) | Up to 58% with optimized PE6 systems [23] | Required 6 optimization strategies for maximal efficiency |
| Large Fragment Insertion | Not supported | Up to 100 bp with 3.12-fold average improvement using EXPERT system [22] | EXPERT enables editing on both sides of nick |
| Zebrafish Embryo Editing | Not reported in search results | 8.4% precise substitution with PE2 vs 4.4% with PEn [24] | PE2 superior for substitutions; PEn better for insertions |
| Off-target Effects | RNA off-target concerns with some deaminases [8] | Minimal reported with proper design [23] | Both show improved specificity over CRISPR-Cas9 |
The performance data demonstrate that base editing typically achieves higher efficiency for its limited set of possible edits, while prime editing offers broader capabilities albeit often with lower initial efficiency. The development of optimized systems like PE5 and PE6, which incorporate dominant-negative MLH1 to suppress mismatch repair, has narrowed this efficiency gap significantly [12] [23]. For therapeutic applications, prime editing has demonstrated remarkable potential in correcting challenging disease-relevant mutations, such as the CFTR F508del variant responsible for cystic fibrosis, achieving correction rates of 25% in patient-derived airway epithelial cells with minimal off-target effects [23].
Experimental Design: Implementation Protocols
Base Editing Workflow
Implementing base editing requires careful consideration of multiple experimental parameters. The following protocol outlines a standard approach for base editing in mammalian cells:
Target Selection and gRNA Design: Identify target sites with the desired base within the editing window (typically positions 4-8 in the protospacer). Avoid sequences with multiple identical bases in the editing window to minimize bystander edits. Verify PAM availability (NGG for SpCas9-based editors) [21].
Editor Selection: Choose the appropriate base editor based on the desired conversion:
- Cytosine Base Editors (CBEs): For C•G to T•A conversions
- Adenine Base Editors (ABEs): For A•T to G•C conversions Consider newer high-fidelity variants to minimize off-target effects [21].
Delivery Method Optimization:
- Plasmid Transfection: Suitable for easily transfectable cell lines
- Viral Delivery: Lentiviral or AAV vectors for challenging cell types
- Ribonucleoprotein (RNP) Complexes: For minimal off-target effects and transient activity
Efficiency Validation:
- Amplicon Sequencing: The gold standard for quantifying editing efficiency
- Restriction Fragment Length Polymorphism: For edits that create or destroy restriction sites
- Sanger Sequencing with Tracking of Indels by Decomposition: For rapid assessment without deep sequencing [21]
Deep learning tools like CRISPRon-ABE and CRISPRon-CBE can significantly improve gRNA design accuracy by predicting editing efficiency and outcome frequencies based on sequence features [21].
Prime Editing Workflow
Prime editing implementation requires more extensive optimization but offers greater versatility. The following protocol outlines key considerations:
pegRNA Design: The most critical factor for successful prime editing:
- Spacer Sequence: Standard 20-nt guide sequence targeting the edit site
- Primer Binding Site (PBS): Typically 10-15 nucleotides, requires optimization for length and complementarity
- Reverse Transcriptase Template (RTT): Encodes the desired edit with sufficient homologous sequence (typically 10-16 nt) beyond the edit site
- 3' Stability Motifs: Incorporate evopreQ1, mpknot, or other RNA motifs to prevent degradation [8] [23]
Editor Selection: Choose the appropriate prime editor variant:
Strategy Optimization:
- PE3 System: Includes an additional nicking guide RNA (ngRNA) targeting the non-edited strand to enhance efficiency through biased repair
- Dual-pegRNA Approaches: For larger edits or replacements
- EXPERT System: Utilizes extended pegRNA with upstream sgRNA for bidirectional editing [22]
Delivery Considerations:
Evaluation Methods:
- Next-generation Sequencing: Essential for comprehensive efficiency assessment
- Edit-to-Indel Ratio Calculation: Critical for evaluating precision
- Off-target Assessment: Whole-genome sequencing for therapeutic applications [23]
Research Reagent Solutions: Essential Tools for Precision Editing
Table 3: Key Reagents for Implementing Precision Genome Editing
| Reagent Category | Specific Examples | Function & Application Notes |
|---|---|---|
| Base Editors | BE4-Gam (CBE), ABE8e (ABE) [21] [23] | Engineered for improved efficiency and specificity; ABE8e shows enhanced activity for A-to-G conversions |
| Prime Editors | PE2, PEmax, PE6 variants [23] [14] | PE6 systems incorporate evolved RT domains and MMR inhibition for enhanced performance |
| Stabilized pegRNAs | epegRNAs [8] [23] | Incorporate 3' RNA motifs (evopreQ1, mpknot) to resist exonuclease degradation |
| Delivery Vectors | AAV, Lentivirus, Lipid Nanoparticles [13] | Dual-AAV systems often required for large prime editor delivery |
| Efficiency Enhancers | MLH1dn, ngRNAs [12] [23] | MLH1dn blocks mismatch repair to improve prime editing outcomes; ngRNAs used in PE3 systems |
| Design Tools | CRISPRon-BE, pegRNA design algorithms [21] | Deep learning models predict editing efficiency and optimize guide design |
Base editing and prime editing represent complementary approaches in the precision genome editing toolkit, each with distinct advantages for specific applications. Base editing offers superior efficiency for straightforward transition mutations within accessible genomic contexts, making it ideal for research applications requiring high efficiency for compatible edits and certain therapeutic contexts where its limitations are not prohibitive [21]. Prime editing provides remarkable versatility for diverse genetic modifications, including transversions, insertions, deletions, and combinations thereof, making it particularly valuable for research requiring precise modeling of human genetic variants and therapeutic applications targeting diverse mutation types [23] [14].
The strategic selection between these technologies depends critically on the specific experimental or therapeutic objectives. For disease modeling and functional genomics where specific point mutations need introduction, base editing often provides the most efficient solution. For therapeutic correction of pathogenic mutations, particularly those involving small insertions or deletions like CFTR F508del, prime editing offers unique capabilities despite historically lower efficiency [23]. As both technologies continue to evolve through protein engineering, guide RNA optimization, and delivery improvements, their complementary strengths will likely expand the frontiers of precision genetic manipulation across basic research and clinical applications.
Practical Deployment in Research and Therapy
The advent of programmable gene editing tools has revolutionized the life sciences, offering unprecedented potential for treating genetic diseases and advancing biological research. Among the current CRISPR-Cas technologies, two primary approaches have emerged for making targeted DNA changes without inducing double-strand breaks: base editing and prime editing [25]. While both technologies represent significant advancements over traditional nuclease-based methods, they differ substantially in their mechanisms, capabilities, and limitations. Base editing, introduced in 2016, enables direct conversion of one DNA base to another through a deamination process but is restricted to specific transition mutations [3] [13]. Prime editing, developed in 2019, provides a more versatile "search-and-replace" mechanism that can install virtually any type of small-scale DNA change without requiring double-strand breaks or donor DNA templates [25] [13]. This article provides a comprehensive comparison of these technologies, focusing on their respective capabilities for installing complex edits, supported by recent experimental data and methodological insights.
Understanding Prime Editing Technology
The Molecular Mechanism of Prime Editing
Prime editing functions as a highly versatile genomic word processor that can precisely rewrite genetic information without causing double-strand DNA breaks [25]. The core prime editing system consists of two main components: a prime editor protein and a specialized prime editing guide RNA (pegRNA) [8] [13]. The editor protein is typically a fusion of a Cas9 nickase (H840A) that cuts only a single DNA strand and an engineered reverse transcriptase (RT) from the Moloney murine leukemia virus [8] [25]. The pegRNA serves a dual purpose: it directs the complex to the target genomic locus through its spacer sequence and encodes the desired edit within its extension region [13].
The editing process occurs through a sophisticated multi-step mechanism [13]. First, the prime editor-pegRNA complex binds to the target DNA site, and the Cas9 nickase creates a single-strand cut in the DNA. The exposed 3' end then hybridizes to the primer binding site (PBS) within the pegRNA, serving as a primer for reverse transcription. The reverse transcriptase then synthesizes DNA using the reverse transcription template (RTT) region of the pegRNA, which contains the desired edit. Finally, cellular repair processes resolve this intermediate structure, incorporating the newly synthesized DNA containing the edit into the genome [25] [13]. This mechanism allows prime editing to achieve a remarkable versatility in genetic modifications while minimizing undesirable byproducts.
Evolution of Prime Editing Systems
Since its initial development, prime editing has undergone significant optimization to improve its efficiency and applicability. The first-generation editor, PE1, established the fundamental architecture but displayed modest editing efficiencies typically below 5% [25]. The introduction of PE2 incorporated engineered mutations to enhance the reverse transcriptase's thermostability, processivity, and binding affinity, resulting in a 1.6- to 5.1-fold increase in editing efficiency compared to PE1 [8] [25]. The PE3 system further improved performance by incorporating an additional sgRNA that nicks the non-edited DNA strand to bias cellular repair toward the edited strand, increasing editing efficiency particularly in challenging genomic contexts [8] [25]. Recent advancements have continued this evolutionary trajectory with systems like PEmax, which incorporates additional optimizations to enhance editing efficiency across diverse targets [19].
Figure 1: The Prime Editing Mechanism. This diagram illustrates the stepwise process of prime editing, from target binding to edit incorporation, highlighting the key molecular events that enable precise genome modification.
Direct Comparison: Prime Editing vs. Base Editing
Editing Scope and Versatility
The most significant distinction between prime editing and base editing lies in their respective editing scopes and versatility. Base editing technology is limited to four specific transition mutations: C-to-T, G-to-A, A-to-G, and T-to-C [3] [13]. This restriction means base editors can only correct approximately 30% of known human pathogenic point mutations [3]. In contrast, prime editing offers substantially broader capabilities, enabling all twelve possible base-to-base conversions, including both transitions and transversions [3] [25]. Additionally, prime editing can efficiently install small insertions and deletions, providing a comprehensive toolkit for precise genome manipulation [8] [25]. Computational models suggest that prime editing could theoretically correct up to 89% of known pathogenic human genetic variants, including single-nucleotide substitutions, small insertions, and deletions [3].
Table 1: Comprehensive Comparison of Editing Capabilities
| Editing Feature | Base Editing | Prime Editing |
|---|---|---|
| Base Substitutions | 4 of 12 possible (C>T, G>A, A>G, T>C) | All 12 possible |
| Insertions | Not supported | Up to dozens of base pairs |
| Deletions | Not supported | Up to dozens of base pairs |
| Theoretical Coverage of Pathogenic Variants | ~30% | Up to 89% |
| Double-Strand Break Formation | No | No |
| Donor DNA Template Required | No | No |
Precision and Editing Purity
Both base editing and prime editing offer advantages over traditional CRISPR-Cas9 nuclease approaches by avoiding double-strand breaks, but they differ significantly in their precision profiles. Base editing operates within an editing window of approximately 4-8 nucleotides and can cause bystander edits where non-target bases within this window are unintentionally modified [3] [6]. For example, adenine base editors (ABEs) exhibit high specificity for A-to-G transitions (97%), while cytosine base editors (CBEs) show slightly lower specificity for C-to-T transitions (92%) [6]. Prime editing generally demonstrates higher precision with minimal bystander editing, as the system directly writes the specified sequence change without deaminating neighboring bases [8]. However, prime editing efficiency can be influenced by cellular mismatch repair mechanisms, which can sometimes reverse installed edits [13]. Recent advancements like the PE5 system address this challenge by incorporating mismatch repair inhibitors to enhance editing persistence [13].
Editing Efficiency and Performance
Editing efficiency represents a crucial practical consideration when choosing between base editing and prime editing technologies. Base editing typically demonstrates higher average efficiency for supported edits, with optimized systems achieving correction rates of 50-90% in many targets [3]. Prime editing efficiency has historically been more variable, ranging from very low single-digit percentages to over 90% depending on the specific edit, target site, and cell type [25] [17]. However, recent optimization efforts have substantially improved prime editing performance. In benchmarked studies using advanced systems like PEmax combined with engineered pegRNAs (epegRNAs) in DNA mismatch repair-deficient cells, prime editing has achieved remarkable efficiencies exceeding 95% for specific edits [19]. These improvements demonstrate the rapid maturation of prime editing technology and its increasing practicality for diverse applications.
Table 2: Experimentally Measured Editing Efficiencies
| Editor Type | Specific Editor | Target | Editing Efficiency | Experimental Conditions |
|---|---|---|---|---|
| Base Editor | ABE7.10 | HEK293T cells | 97% A>G specificity | SURRO-seq data, ~11,500 gRNAs [6] |
| Base Editor | BE4-Gam | HEK293T cells | 92% C>T specificity | SURRO-seq data, ~11,500 gRNAs [6] |
| Prime Editor | PEmax + epegRNA | HEK3 +1 T>A | 95% precise editing | MMR-deficient cells, 28 days [19] |
| Prime Editor | PEmax + epegRNA | DNMT1 +6 G>C | 95% precise editing | MMR-deficient cells, 28 days [19] |
| Prime Editor | PE2 | Various plant species | Highly variable (0.0%-29.2%) | Across species, targets, and edit types [17] |
Experimental Data and Benchmarking
Methodologies for Assessing Editing Performance
Rigorous assessment of editing technologies requires carefully designed experimental protocols and benchmarking strategies. Recent high-quality studies have employed sophisticated methodologies to quantitatively evaluate base and prime editing performance. For base editing, the SURRO-seq technology has enabled large-scale quantification of editing efficiency and outcomes [6]. This approach involves creating lentiviral gRNA-target pair libraries with approximately 1,000x coverage, transducing editor-expressing cells (e.g., HEK293T-ABE or HEK293T-CBE) at low multiplicity of infection (MOI=0.3), and harvesting cells after 8 days of growth under selective conditions [6]. Deep amplicon sequencing of surrogate target site-specific PCR products then provides comprehensive data on editing efficiencies and outcomes.
For prime editing assessment, recent benchmark studies have utilized stable cell lines constitutively expressing editor proteins (PE2 or PEmax) combined with self-targeting sensor libraries that link epegRNA expression cassettes to target sequences [19]. These systems allow editing to be analyzed across thousands of guide RNA-target pairs while modeling endogenous target editing. In typical experiments, libraries are transduced at low MOI, cells are selected for cassette integration, and populations are grown for extended periods (e.g., 28 days) with periodic sampling to track editing progression [19]. Sequencing data are then analyzed to quantify precise edits, errors, and unedited sequences, providing a comprehensive view of editing performance across diverse targets.
Recent Performance Breakthroughs
Substantial progress has been made in addressing the historical limitation of variable prime editing efficiency. Research published in Nature Methods in 2024 demonstrated that combining stable editor expression with MMR deficiency and optimized epegRNAs enables remarkably high editing efficiencies [19]. In these optimized conditions, prime editing achieved up to 95% precise editing at endogenous targets like HEK3 and DNMT1, with minimal co-occurrence of unwanted outcomes [19]. The study further established that precise editing accumulates over time with stable editor expression, ultimately reaching near-saturation levels for many targets by day 28 in culture [19].
Large-scale screening approaches have further demonstrated the capabilities of optimized prime editing systems. A platform employing approximately 240,000 engineered epegRNAs targeting ~17,000 codons successfully identified negative selection phenotypes for 7,996 nonsense mutations across 1,149 essential genes [19]. This achievement highlights prime editing's capacity for multiplexed functional characterization of genetic variants with simple readouts, establishing its utility for high-throughput genetic screening applications.
Research Reagent Solutions
Table 3: Essential Research Reagents for Prime Editing Experiments
| Reagent Category | Specific Examples | Function and Importance |
|---|---|---|
| Editor Proteins | PE2, PEmax, PE3, PE5 | Core editing machinery combining nCas9 with reverse transcriptase; optimized versions enhance efficiency and specificity [8] [19] |
| Guide RNAs | pegRNA, epegRNA | Specify target site and encode desired edit; engineered versions (e.g., with tevopreQ1 motif) improve stability and efficiency [8] [19] |
| Delivery Systems | AAV vectors, lipid nanoparticles (LNPs), electroporation | Enable intracellular delivery of editing components; dual-vector systems often required due to large size of editor constructs [3] [13] |
| Cell Engineering Tools | MLH1 knockout, MMR inhibitors | Suppress mismatch repair to enhance editing persistence; crucial for achieving high efficiency with small edits [13] [19] |
| Design Tools | CRISPRon-ABE/CBE, pegRNA design algorithms | Computational tools for predicting editing efficiency and optimizing guide RNA design; essential for experimental success [6] [26] |
Figure 2: Prime Editing Experimental Workflow. This diagram outlines the key stages of a prime editing experiment, from initial design to final analysis, highlighting critical optimization points that influence editing success.
Prime editing represents a transformative advancement in precision genome editing technology, offering unprecedented versatility for installing complex genetic modifications. While base editing remains a powerful tool for specific transition mutations, prime editing's ability to perform all possible base substitutions, insertions, and deletions positions it as the more comprehensive solution for precise genome manipulation. Recent experimental breakthroughs have addressed earlier limitations in editing efficiency, with optimized systems now achieving >95% precision in benchmarked targets [19]. The development of sophisticated design tools [6] [26] and delivery solutions [3] [13] further enhances prime editing's practicality for diverse research applications. As the technology continues to evolve, prime editing is poised to enable new frontiers in functional genomics, disease modeling, and therapeutic development, providing researchers with an increasingly powerful toolkit for precise genetic engineering.
Functional genomics aims to understand the relationship between genetic sequence and biological function. A central challenge in this field is the interpretation of the vast number of genetic variants discovered through sequencing, most of which are classified as variants of uncertain significance (VUS) [27]. For clinicians and researchers, determining which of these variants contribute to disease is crucial for risk assessment and developing therapeutic strategies.
Traditional methods for characterizing variants have faced significant limitations. CRISPR-Cas9 nuclease-based approaches rely on double-strand breaks (DSBs), leading to unpredictable insertions and deletions (indels) that complicate the interpretation of phenotypic outcomes [8] [9]. While base editing offered a more precise solution without DSBs, its application is restricted, as it can only mediate four of the twelve possible base-to-base conversions and is prone to unwanted bystander edits within its activity window [3] [13].
The emergence of prime editing has revolutionized this landscape. As a versatile "search-and-replace" technology, it enables all 12 base-to-base conversions, as well as small insertions and deletions, without inducing DSBs [8] [13]. This precision has made it an ideal tool for saturation screening, a powerful approach where hundreds to thousands of genetic variants are introduced and functionally characterized in a single, high-throughput experiment [28] [27]. This guide objectively compares how prime editing is being applied in functional genomics screens against the capabilities of base editing, detailing the experimental protocols, key findings, and essential reagents that define this cutting-edge methodology.
Technology Comparison: Prime Editing vs. Base Editing
At its core, prime editing systems consist of a prime editor protein and a specialized prime editing guide RNA (pegRNA). The editor protein is a fusion of a Cas9 nickase (H840A) and an engineered reverse transcriptase (RT). The pegRNA not only specifies the target site but also contains a template for the desired edit [8] [13]. The process involves nicking the target DNA, hybridizing a primer binding site (PBS), and reverse-transcribing the new genetic information from the pegRNA template into the genome [13].
Base editors, in contrast, use a catalytically impaired Cas9 (nickase) fused to a deaminase enzyme. Cytosine base editors (CBEs) convert C•G to T•A base pairs, while adenine base editors (ABEs) convert A•T to G•C [9]. They do not create DSBs but achieve only a limited subset of possible nucleotide changes and can cause unintended edits at nearby bases, a phenomenon known as bystander editing [3] [21].
Table 1: Fundamental Comparison of Prime Editing and Base Editing Technologies
| Feature | Prime Editing | Base Editing |
|---|---|---|
| Core Components | Cas9 nickase (H840A) + Reverse Transcriptase + pegRNA | Cas9 nickase + Deaminase Enzyme (e.g., APOBEC1, TadA) + sgRNA |
| DNA Cleavage | Single-strand nick (no DSBs) | Single-strand nick or no cleavage (no DSBs) |
| Editing Scope | All 12 base substitutions, insertions, deletions | Primarily C>G to T>A (CBEs) and A>T to G>C (ABEs) |
| Theoretical Coverage | ~89% of known pathogenic human genetic variants [3] | Limited to specific transition mutations |
| Key Limitation | Large size complicates delivery; variable efficiency | Bystander edits; restricted to specific base transitions |
| Ideal Application | Saturation editing of all variant types | High-throughput screening of specific transition mutations |
Saturation Prime Editing (SPE) in Practice: Experimental Data and Protocols
Saturation Prime Editing (SPE) leverages the precision of prime editing to systematically introduce thousands of genetic variants into a genomic locus, enabling their functional impact to be assessed in a single, pooled experiment.
Key Applications and Findings
- Functional Characterization of NPC1 and BRCA2: Researchers applied SPE to the NPC1 gene, mutations in which cause Niemann-Pick disease type C. The screen assayed 706 missense mutations, classifying 410 as deleterious, indicating the gene's high sensitivity to genetic perturbation. The method was also successfully translated to the BRCA2 gene, demonstrating its generalizability for genes with appropriate cellular assays [28].
- Mass Screening of Tumor Suppressor Genes: At the Francis Crick Institute, researchers used pooled prime editing to screen variants in the tumor suppressor genes SMARCB1 and MLH1 (a risk gene for bowel cancer). For MLH1, they scored over 700 variants, newly identifying loss-of-function variants in both coding and non-coding regions. This approach has the potential to screen thousands of variants in a single experiment, clarifying disease risk for many VUS [27].
- Non-Coding Variant Analysis with PRIME: A method dubbed PRIME was developed for pooled prime editing screens, enabling the characterization of thousands of coding and non-coding variants. In one application, researchers performed saturation mutagenesis on a 716 bp MYC enhancer. They designed a library of 6,252 pegRNA/ngRNA pairs to generate 2,148 single-nucleotide substitutions, successfully identifying essential nucleotides for the enhancer's function [29].
Table 2: Summary of Key Saturation Prime Editing Studies
| Study Focus | Gene/Locus | Scale of Screening | Key Outcome |
|---|---|---|---|
| Variant Interpretation [28] | NPC1, BRCA2 | 706 missense mutations in NPC1 | 410 (58%) of assayed NPC1 variants classified as deleterious |
| Tumor Suppressor Gene Screening [27] | MLH1, SMARCB1 | >700 variants in MLH1 | Identified a cluster of loss-of-function variants in MLH1 |
| Enhancer Mapping [29] | MYC Enhancer | 2,148 single-nucleotide substitutions | Identified essential nucleotides for a 716 bp enhancer's function |
Detailed Experimental Protocol: A Workflow for Pooled SPE
The following diagram illustrates a generalized workflow for a pooled saturation prime editing screen, integrating common steps from the cited studies.
Step-by-Step Protocol:
- pegRNA Library Design and Cloning: A library of pegRNAs is designed to introduce the desired set of substitutions, insertions, or deletions across the target genomic region. Each pegRNA includes a spacer sequence for targeting, a primer binding site (PBS), and a reverse transcriptase template (RTT) encoding the edit. To enhance efficiency, structured RNA motifs (e.g., evopreQ1) are often added to the 3' end of the pegRNA to prevent degradation [8] [29]. This library is then cloned into a lentiviral backbone.
- Lentiviral Library Production: The pooled plasmid library is used to produce a lentiviral library. The titer is carefully controlled to ensure a low multiplicity of infection (MOI ~0.3), guaranteeing that most cells receive only one viral particle and thus one pegRNA [29] [21].
- Cell Infection and Selection: Target cells—often engineered to stably express the prime editor protein (nCas9-RT fusion)—are infected with the lentiviral pegRNA library. This stable expression system, as used in the "PRIME" protocol, significantly improves editing efficiency over co-delivery methods [29]. Cells are then selected with antibiotics (e.g., puromycin) to generate a pooled population of edited cells.
- Phenotypic Selection and Expansion: The cell pool is cultured for a period sufficient for a phenotypic outcome to manifest. For studies identifying essential genes or functional domains, this involves passaging the cells for several weeks. Cells with edits that compromise function (e.g., in a tumor suppressor gene) are depleted over time, while those with neutral edits remain [29] [27].
- Sequencing and Data Acquisition: Genomic DNA is harvested at an initial time point (e.g., Day 2 post-selection) and after phenotypic selection (e.g., Day 30). The integrated pegRNA sequences are amplified by PCR and quantified by next-generation sequencing to track the abundance of each pegRNA over time [28] [29].
- Data Analysis: The fold-change in abundance for each pegRNA between the initial and final time points is calculated using specialized pipelines like MAGeCK. pegRNAs that are significantly depleted are inferred to target functionally essential residues or regions [29].
The Scientist's Toolkit: Essential Reagents for SPE
Table 3: Key Research Reagent Solutions for Saturation Prime Editing
| Reagent / Solution | Function in SPE | Key Considerations |
|---|---|---|
| Prime Editor Proteins | Catalyzes the nick and reverse transcription steps. Common versions include PE2 and PE3. | PE2 offers a balance of efficiency and simplicity; PE3 uses a second nicking gRNA to boost efficiency but may slightly increase indel rates [8] [13]. |
| Structured pegRNAs (epegRNAs) | Guides the editor to the target and templates the edit; engineered with stabilizing motifs. | Motifs like evopreQ1 or mpknot at the 3' end protect against exonucleases, improving editing efficiency 3-4 fold [8] [29]. |
| Lentiviral Delivery Systems | Enables efficient, stable delivery of the pegRNA library into a pooled cell population. | Critical for achieving low MOI to ensure one edit per cell. Requires high-diversity library production [29] [27]. |
| Stable Cell Lines | Mammalian cells (e.g., MCF7, HEK293T) engineered to constitutively express the prime editor protein. | Simplifies screening by requiring only delivery of the pegRNA library, leading to higher and more consistent editing efficiencies [29]. |
| Deep Sequencing Platforms | Quantifies the abundance of each pegRNA in the population before and after phenotypic selection. | Essential for generating quantitative data on variant function. Requires high coverage and accuracy [28] [29]. |
Saturation prime editing represents a significant leap forward in functional genomics. By enabling the precise, high-throughput functional assessment of nearly any genetic variant—including single-nucleotide changes, indels, and non-coding mutations—SPE is overcoming the major limitations of previous technologies like DSB-dependent CRISPR/Cas9 and the restricted scope of base editing [28] [27]. As methods for optimizing pegRNA design, editor efficiency, and in vivo delivery continue to advance, the capacity of SPE to map genetic function at base-pair resolution will be critical for realizing the promise of personalized medicine, from accurate disease risk prediction to the development of novel, genetically-informed therapies.
The field of crop improvement is undergoing a revolutionary transformation with the advent of precision genome editing technologies. While traditional CRISPR-Cas9 systems have provided powerful tools for functional genomics, their reliance on creating double-strand breaks (DSBs) in DNA often leads to unpredictable editing outcomes, including insertions, deletions, and complex rearrangements [8] [9]. These limitations have prompted the development of more precise genetic modification tools, notably base editing and prime editing, which enable targeted nucleotide changes without inducing DSBs or requiring donor DNA templates [3] [13]. For researchers and scientists focused on crop enhancement, these advanced editing platforms offer unprecedented opportunities to develop improved varieties with precise genetic modifications that mimic naturally occurring alleles, potentially streamlining regulatory approval processes [30].
Base editing and prime editing represent complementary approaches in the precision breeding toolbox, each with distinct mechanisms, capabilities, and limitations. Base editing, described as "CRISPR 2.0," utilizes a catalytically impaired Cas protein fused to a nucleobase deaminase enzyme to directly convert one DNA base to another without breaking the DNA backbone [31] [9]. Prime editing, often characterized as a "search-and-replace" system, employs a Cas9 nickase fused to a reverse transcriptase that is programmed with an extended guide RNA (pegRNA) to copy edited genetic information directly into the target DNA site [8] [13]. Understanding the technical specifications, performance parameters, and optimal applications of each system is essential for plant biotechnologists seeking to address different breeding challenges, from correcting single nucleotide polymorphisms (SNPs) to introducing small gene insertions or deletions that can enhance crop productivity, nutritional quality, and stress resilience.
Molecular Mechanisms and System Architectures
Base Editing Architecture and Editing Mechanism
Base editors are sophisticated molecular machines that enable the direct conversion of one DNA base pair to another without requiring DSBs. These systems consist of three core components: a catalytically impaired Cas nuclease (typically a nickase variant that cuts only one DNA strand), a nucleobase deaminase enzyme, and in some configurations, an inhibitor of base excision repair [3] [9]. The most established base editor classes are Cytosine Base Editors (CBEs), which convert C•G base pairs to T•A, and Adenine Base Editors (ABEs), which convert A•T base pairs to G•C [9]. These four transition mutations (C-to-T, G-to-A, A-to-G, T-to-C) account for approximately 60% of all known pathogenic single nucleotide variants in humans, suggesting similar coverage for agriculturally important alleles [3] [9].
The base editing process initiates when the guide RNA directs the base editor complex to the target DNA sequence. The Cas component partially unwinds the DNA, exposing a single-stranded region known as the "editing window" (typically 4-5 nucleotides in width) where the deaminase enzyme acts [8]. Following deamination, the edited strand is preferentially nicked by the Cas nickase, encouraging the cell's repair machinery to replace the unedited strand using the edited strand as a template, thereby permanently installing the desired base change [13]. While this mechanism avoids DSBs, a significant limitation of base editors is "bystander editing," where additional bases within the editing window are unintentionally modified, potentially leading to undesirable outcomes [3]. Recent engineering efforts have focused on narrowing the editing window and developing high-fidelity deaminase variants to mitigate this issue.
Prime Editing Architecture and Editing Mechanism
Prime editing represents a more versatile but complex approach to precision genome editing. The system comprises two fundamental components: (1) a prime editor protein, which is a fusion of a Cas9 nickase (H840A) with an engineered reverse transcriptase (RT) from the Moloney Murine Leukemia Virus (MMLV), and (2) a prime editing guide RNA (pegRNA) that both specifies the target site and encodes the desired edit [8] [13]. Unlike standard guide RNAs, pegRNAs contain a 3' extension consisting of a primer binding site (PBS) and a reverse transcriptase template (RTT) that includes the desired genetic modification [13].
The prime editing mechanism occurs through a multi-step process that begins with the pegRNA-directed binding of the prime editor to the target DNA site. The Cas9 nickase then creates a single-strand cut in the non-target DNA strand, exposing a 3'-hydroxyl group that serves as a primer for the RT [8] [13]. This primer hybridizes to the PBS sequence of the pegRNA, enabling the RT to synthesize a DNA flap using the RTT as a template. The newly synthesized edited flap then competes with the original unedited flap for incorporation into the genome through cellular DNA repair processes [13]. To increase editing efficiency, particularly in challenging genomic contexts, additional systems like PE3 incorporate a second standard sgRNA that nicks the non-edited strand, encouraging the cell to use the edited strand as a repair template [8] [12]. The requirement for multiple hybridization events contributes to prime editing's high specificity but can also limit its efficiency in some applications.
Figure 1: Prime Editing Molecular Mechanism. The process involves four key steps: (1) target recognition and binding of the PE:pegRNA complex to DNA, (2) nicking of the non-target DNA strand, (3) reverse transcription using the pegRNA template, and (4) resolution and integration of the edited DNA flap into the genome [8] [13].
Comparative Performance Analysis
Technical Specifications and Editing Capabilities
The selection between base editing and prime editing for a specific crop improvement application requires careful consideration of their fundamental technical capabilities. Base editors offer a more limited editing scope but generally higher efficiency for the specific transitions they catalyze, while prime editors provide substantially broader editing capabilities but often with more variable efficiency that depends heavily on target context and optimization [8] [3] [13].
Table 1: Technical Capabilities Comparison Between Base Editing and Prime Editing
| Editing Feature | Base Editing | Prime Editing |
|---|---|---|
| DSB Formation | No | No |
| Donor DNA Required | No | No |
| Point Mutations | 4 transitions (C>T, G>A, A>G, T>C) | All 12 possible point mutations |
| Small Insertions | No | Yes (typically <50 bp) |
| Small Deletions | No | Yes (typically <50 bp) |
| Theoretical Correction Rate* | ~30% of known pathogenic SNPs | ~89% of known pathogenic SNPs |
| Key Limitation | Bystander edits, PAM dependency | Variable efficiency, complex pegRNA design |
| Ideal Use Case | Straightforward transition mutations | Complex edits including transversions, insertions, deletions |
Note: Theoretical correction rates are extrapolated from human pathogenic mutations and illustrate relative coverage potential [3].
Efficiency and Specificity Metrics
Quantitative assessment of editing performance reveals significant differences between these platforms. Base editors typically achieve higher editing efficiencies (often 30-70% in mammalian cells) for their compatible substitutions but suffer from bystander editing where adjacent bases within the editing window are unintentionally modified [8] [3]. Prime editing efficiencies vary more widely (typically 10-50% in mammalian cells without optimization) but continue to improve with system engineering [8] [12]. Both systems exhibit substantially fewer off-target effects compared to conventional CRISPR-Cas9 nucleases, with prime editing demonstrating particularly high specificity due to its requirement for three separate hybridization events for successful editing [8] [13].
Table 2: Experimental Performance Metrics in Model Systems
| Performance Parameter | Base Editing | Prime Editing | Notes |
|---|---|---|---|
| Typical Editing Efficiency | 30-70% | 10-50% (highly variable) | Efficiency depends on cell type, delivery method, and target locus [8] [12] |
| Indel Formation | <1% | 0.5-5.0% | Significantly lower than DSB-based methods [8] |
| Bystander Editing | Yes (significant concern) | Minimal | Bystander edits occur at non-target bases within editing window [3] |
| Off-Target Effects | Low (RNA-dependent) | Very low | Both substantially better than CRISPR-Cas9 nucleases [8] |
| PAM Flexibility | Limited to Cas variant used | Limited to Cas variant used | Both systems benefit from emerging Cas variants with relaxed PAM requirements [32] |
Recent advancements have substantially improved prime editing efficiency. The development of engineered pegRNAs (epegRNAs) with structured RNA motifs at their 3' terminus protects against degradation and improves editing efficiency by 3-4 fold across multiple cell types [8]. Systems like PE4 and PE5 incorporate dominant-negative MLH1 (MLH1dn) to suppress mismatch repair, increasing efficiency to 50-80% in some contexts [12]. The innovative proPE (prime editing with prolonged editing window) system uses a second non-cleaving sgRNA to target the reverse transcriptase template near the edit site, increasing editing efficiency up to 29.3% for previously low-performing edits (<5% with standard PE) and expanding the editable range [32].
Experimental Implementation in Crop Systems
Research Reagent Solutions Toolkit
Successful implementation of base editing or prime editing in crop species requires careful selection and optimization of molecular tools. The following research reagent solutions represent essential components for designing precision genome editing experiments in plants.
Table 3: Essential Research Reagents for Precision Genome Editing in Plants
| Reagent Category | Specific Examples | Function & Application Notes |
|---|---|---|
| Editor Proteins | BE4max, ABE8e, PE2, PEmax, PE5 | Engineered for improved efficiency and specificity; codon-optimize for plant species [8] [12] |
| Guide RNA Systems | pegRNAs, epegRNAs, sgRNAs | pegRNA design critical for prime editing success; epegRNAs improve stability [8] |
| Delivery Vehicles | Agrobacterium T-DNA, gold particle bombardment, viral vectors (e.g., Bean Yellow Dwarf Virus), lipid nanoparticles | Plant-specific delivery constraints; consider cargo size limitations especially for prime editors [8] [30] |
| Selectable Markers | Fluorescent proteins, antibiotic resistance genes, metabolic selection | Enable identification and isolation of successfully edited plant cells during regeneration |
| Validation Tools | Sanger sequencing, amplicon deep sequencing, restriction fragment length analysis, T7E1 assay | Essential for confirming intended edits and detecting potential off-target effects |
Optimized Experimental Protocols
Implementing precision editing in crop species requires adaptation of standard protocols to address plant-specific challenges such as cell wall barriers, regeneration efficiency, and the frequent need for elimination of editor components from final edited lines. The following protocols summarize key methodologies for successful base editing and prime editing in plant systems.
Protocol 1: Prime Editing Implementation in Plants
- Target Selection & pegRNA Design: Identify target site with appropriate PAM sequence (typically SpCas9 NGG). Design pegRNA with 10-15 nt primer binding site (PBS) and reverse transcriptase template (RTT) containing desired edit with 2-15 nt homologous flanking sequence. Consider using epegRNA designs with evopreQ or mpknot motifs for improved stability [8].
- Vector Construction: Clone pegRNA expression cassette (typically under U6/U3 pol III promoter) and prime editor expression cassette (under pol II promoter) into plant transformation vector. For challenging edits, consider co-expressing MLH1dn to inhibit mismatch repair [12].
- Plant Transformation: Deliver constructs using Agrobacterium-mediated transformation (for dicots) or particle bombardment (for monocots). Include appropriate selectable markers for transgenic tissue identification.
- Regeneration & Selection: Regenerate whole plants from transformed tissue under selection pressure. Multiple regeneration cycles may be needed to obtain homogeneously edited plants.
- Molecular Validation: Genotype regenerated plants by PCR and sequencing across target locus. Perform amplicon deep sequencing to quantify editing efficiency and detect potential byproducts. Screen for presence of editor components in final lines.
Protocol 2: Base Editing Optimization in Plants
- Target Selection & gRNA Design: Identify target site with desired base change positioned within the editing window (typically positions 4-8 for SpCas9-based editors). Avoid sites with multiple identical bases in editing window to minimize bystander edits [3].
- Base Editor Selection: Choose appropriate cytosine (CBE) or adenine (ABE) base editor based on desired base conversion. Consider high-fidelity variants to minimize off-target editing.
- Delivery & Regeneration: Similar to Protocol 1, though base editors' smaller size may enable delivery options not feasible for prime editors.
- Editing Assessment: Sequence target locus to confirm intended base changes. Quantify efficiency and screen extensively for bystander edits at adjacent positions within the editing window. For crops, evaluate editing uniformity across tissues.
Figure 2: Decision Framework for Editor Selection. This workflow guides researchers in selecting the appropriate precision editing technology based on the specific genetic modification objective, highlighting the complementary applications of base editing and prime editing [8] [3] [13].
Applications in Crop Improvement
Precision genome editing technologies are being deployed to address numerous challenges in crop production and quality. Unlike conventional genetic engineering approaches that often introduce foreign DNA, base editing and prime editing can potentially create improvements that are indistinguishable from naturally occurring mutations, offering a regulatory path that may accelerate commercialization [30].
Base editing has proven particularly valuable for introducing single nucleotide changes that confer important traits. Prime editing's versatility makes it suitable for more complex modifications, including:
- Herbicide tolerance: Creating targeted amino acid substitutions in herbicide target proteins that confer resistance while maintaining normal enzymatic function [30].
- Disease resistance: Engineering resistance alleles by modifying susceptibility genes involved in pathogen recognition or defense signaling [30].
- Nutritional enhancement: Optimizing protein quality through targeted changes in storage protein genes to improve amino acid balance [30].
- Yield components: Fine-tuning regulatory sequences or coding regions of genes controlling grain size, panicle architecture, or fruit development [30].
- Environmental resilience: Introducing protective alleles that enhance drought tolerance, nitrogen use efficiency, or thermal adaptability [30].
Notable success stories include the development of CRISPR-edited cassava with resistance to viral diseases that threaten food security across Africa [30]. Similarly, climate-resilient wheat and rice varieties capable of withstanding extreme drought, heat, and flooding have been developed using precision editing approaches [30]. The nitrogen-efficient legumes and cereals created through targeted genome modifications represent another significant advancement, potentially reducing synthetic fertilizer requirements by up to 30% while maintaining yield [30].
Base editing and prime editing represent complementary pillars in the expanding toolkit of precision breeding technologies. While base editing offers higher efficiency for straightforward transition mutations, prime editing provides unprecedented versatility for diverse genetic modifications including transversions, insertions, and deletions. The ongoing optimization of both systems—through improved editor proteins, stabilized guide RNA designs, and enhanced delivery methods—continues to expand their applicability across crop species.
For crop researchers and developers, the strategic selection between these technologies depends on multiple factors: the specific genetic change required, the efficiency threshold needed for practical application, regulatory considerations, and the resources available for optimization. Base editing may be preferable for simple transition mutations where high efficiency is paramount, while prime editing becomes essential for more complex edits beyond the scope of base editors. Emerging variants like PE5, PE6, and PE7 with efficiency reaching 80-95% in mammalian cells suggest a trajectory of continuous improvement that will further enhance their utility in plant systems [12].
The future of precision crop improvement will likely see increased use of these technologies in combination, leveraging the strengths of each platform to pyramid multiple desirable traits. As regulatory frameworks evolve to distinguish these precise editing approaches from traditional transgenic methods, and as delivery efficiency improves across diverse crop species, base editing and prime editing are poised to become fundamental tools for developing the next generation of improved crop varieties that will support sustainable agricultural systems and global food security.
Navigating Challenges and Enhancing Editor Performance
Addressing Off-Target and Bystander Editing in Base Editors
The advent of CRISPR-based base editing has revolutionized precision genome engineering by enabling direct chemical conversion of one DNA base into another without inducing double-strand breaks (DSBs). This approach represents a significant advancement over conventional CRISPR-Cas9 nucleases, which rely on creating DSBs that can lead to unpredictable insertions, deletions, and chromosomal rearrangements. Base editors have emerged as powerful tools for correcting point mutations that account for a substantial proportion of known genetic disorders. However, as these technologies have advanced toward therapeutic applications, two significant challenges have emerged: off-target editing at unintended genomic locations and bystander editing at non-target nucleotides within the editing window. These precision limitations have prompted rigorous comparison with the newer prime editing technology, which offers a fundamentally different mechanism for genomic correction. Understanding the relative strengths and limitations of each system is crucial for researchers and therapeutic developers selecting the appropriate precision editing tool for specific applications. This review comprehensively compares the off-target and bystander editing profiles of base editors against prime editors, providing experimental data and methodologies essential for informed platform selection in research and therapeutic development.
Molecular Mechanisms: Architectural Differences Underlying Precision
The divergent precision profiles of base editors and prime editors stem from their fundamentally different molecular architectures and editing mechanisms. Understanding these core mechanisms is essential for interpreting their performance differences in off-target and bystander editing.
Base Editor Architecture and Deamination Mechanism
Base editors are modular fusion proteins consisting of a catalytically impaired Cas9 nickase (nCas9) tethered to a nucleobase deaminase enzyme. The system functions through a deamination mechanism that directly converts one base to another without DNA backbone cleavage. Two primary classes of base editors have been developed: cytosine base editors (CBEs) that convert cytosine to thymine (C-to-T) through a uracil intermediate, and adenine base editors (ABEs) that convert adenine to guanine (A-to-G) through an inosine intermediate. CBEs typically incorporate a cytidine deaminase (such as APOBEC1) along with a uracil glycosylase inhibitor (UGI) to prevent uracil excision and improve editing efficiency. ABEs utilize an engineered tRNA-specific adenosine deaminase (TadA) that has been evolved to act on single-stranded DNA. Both systems operate within a defined editing window of approximately 4-10 nucleotides, where the deaminase enzyme can access exposed single-stranded DNA generated by the nCas9 binding to the target site. This editing window represents a critical limitation, as any editable bases within this region are susceptible to modification, leading to bystander edits [12] [33] [9].
Figure 1: Comparative mechanisms of base editing and prime editing. Base editors (top) can modify multiple nucleotides within an editing window, while prime editors (bottom) incorporate only the specifically programmed edit.
Prime Editor Architecture and Search-and-Replace Mechanism
Prime editors employ a fundamentally different "search-and-replace" mechanism that avoids the limitations of deaminase-based editing. The system comprises a nCas9 (H840A) domain fused to an engineered reverse transcriptase (RT) domain, programmed with a specialized prime editing guide RNA (pegRNA). The pegRNA serves dual functions: it directs the complex to the target genomic locus through its spacer sequence and encodes the desired edit within its reverse transcriptase template (RTT) region. Upon binding to the target DNA, the nCas9 nicks one strand, creating a primer for the reverse transcriptase to synthesize DNA containing the desired edit using the pegRNA as a template. This mechanism enables precise installation of all 12 possible base-to-base conversions, small insertions, and deletions without DSBs. The requirement for multiple hybridization events between the pegRNA and target DNA enhances specificity, while the absence of deaminase enzymes eliminates the bystander editing characteristic of base editors. The original PE system has evolved through several generations (PE1-PE6), with each iteration improving editing efficiency through protein engineering and optimization of accessory components [12] [13] [8].
Quantitative Comparison: Off-Target and Bystander Editing Profiles
Rigorous comparative studies have quantified the substantial differences in editing precision between base editing and prime editing platforms. The data reveal critical trade-offs between editing efficiency and precision that inform platform selection for specific applications.
Table 1: Comparative Performance of Base Editors and Prime Editors
| Editing Characteristic | Cytosine Base Editors (CBEs) | Adenine Base Editors (ABEs) | Prime Editors |
|---|---|---|---|
| Primary Editing Mechanism | Chemical deamination of C to U | Chemical deamination of A to I | Reverse transcription from RNA template |
| Editing Window | 4-10 nucleotides | 4-10 nucleotides | Programmatically determined |
| Bystander Editing | Frequent within editing window | Frequent within editing window | Minimal to none |
| Off-Target DNA Editing | Cas9-dependent and deaminase-dependent | Cas9-dependent and deaminase-dependent | Primarily Cas9-dependent |
| Off-Target RNA Editing | Significant with some variants | Lower than CBEs | Minimal |
| Typical Efficiency Range | 10-70% (highly target-dependent) | 10-60% (highly target-dependent) | 5-50% (highly optimized systems can reach >90%) |
| Indel Formation | Low (<1-5%) | Low (<1-5%) | Very low (<1%) |
| Therapeutic Versatility | Limited to C→T, G→A transitions | Limited to A→G, T→C transitions | All 12 possible base conversions, insertions, deletions |
Bystander Editing Experimental Evidence
Bystander editing represents a particularly challenging limitation for base editors, as multiple nucleotides within the editing window can be modified simultaneously with the intended target base. A compelling demonstration of the functional consequences of bystander editing comes from a 2024 study evaluating ABE-mediated correction of the Rpe65 mutation in a mouse model of Leber congenital amaurosis (LCA). Researchers compared three ABE variants (ABEmax, ABE8e, and ABE8eWQ) for their ability to correct the disease-causing nonsense mutation (c.130C>T, p.R44X) while monitoring bystander editing at adjacent adenines [34].
The study revealed that ABE8e, which has the highest on-target editing activity, also generated substantial bystander editing at positions A3, A8, and A11 (counting the end distal to the PAM as position 1). Specifically, ABE8e produced average bystander editing rates of 12.32% at A3, 14.77% at A8, and 5.49% at A11 in retinal tissue, alongside 16.38% on-target editing at the pathogenic A6 position. Critically, the L43P mutation resulting from bystander editing at A3 was predicted through AlphaFold-based mutational scanning and molecular dynamics simulations to disrupt RPE65 protein structure and function. Despite successful correction of the primary mutation and RPE65 expression restoration, visual function failed to recover in ABE8e-treated mice due to these deleterious bystander edits [34].
In contrast, prime editors introduced in the same study achieved precise correction of the pathogenic mutation without bystander edits, demonstrating the critical advantage of prime editing for therapeutic applications where collateral editing could compromise functional outcomes. The requirement for multiple hybridization events in prime editing - target binding, primer binding site annealing, and template hybridization - creates a built-in specificity mechanism that minimizes off-target effects while virtually eliminating bystander editing [12] [8].
Experimental Approaches for Assessing Editing Precision
Robust assessment of editing precision requires standardized experimental protocols that comprehensively evaluate both on-target and off-target editing. The following methodologies represent current best practices for characterizing base editor and prime editor specificity.
Bystander Editing Quantification Protocol
The rd12 mouse model study established a comprehensive protocol for evaluating bystander editing in vivo:
Dual AAV Vector Delivery: Package base editor components (TadA-8e or other variants with NG-Cas9 nickase) into separate AAV vectors using a split-intein system to accommodate size constraints of AAV packaging. Administer via subretinal injection to 3-week-old rd12 mice [34].
Temporal Analysis: Allow 6 weeks for editing and expression, then harvest retinal tissue for parallel genotyping and functional assessment.
High-Throughput Sequencing: Amplify target genomic regions using primers flanking the edit site. Perform deep sequencing (>10,000x coverage) to detect low-frequency editing events.
Bioinformatic Analysis: Quantify editing percentages at each position within the editing window using tools like CRISPResso2. Normalize frequencies to total reads.
Functional Correlation: Assess protein expression (e.g., Western blot for RPE65) and functional recovery (electroretinography for visual function) to correlate bystander edits with phenotypic outcomes.
This integrated approach enables researchers to not only quantify bystander editing frequencies but also determine their functional significance, which is crucial for therapeutic development [34].
Off-Target Editing Assessment Methods
Multiple methodologies have been developed to comprehensively profile off-target editing by base editors and prime editors:
Guide RNA-Independent Off-Target Screening:
- CELL-seq: Captures transcriptome-wide RNA off-target editing by base editor deaminases, particularly important for CBEs which can exhibit significant RNA editing [12].
- R-loop Assays: Evaluate deaminase activity on transiently exposed single-stranded DNA during transcription, identifying potential off-target sites independent of Cas9 binding.
Cas9-Dependent Off-Target Detection:
- GUIDE-seq: Uses tagged double-stranded oligonucleotides to integrate into DSB sites, comprehensively mapping Cas9-dependent off-target sites across the genome.
- Circle-seq: In vitro approach that uses circularized genomic DNA to identify potential Cas9 cleavage sites without cellular context limitations.
Computational Prediction and Validation:
- In silico prediction: Tools like Cas-OFFinder predict potential off-target sites based on sequence similarity to the target site.
- Targeted amplicon sequencing: Deep sequencing of predicted off-target loci to quantify editing frequencies.
Prime editors generally demonstrate superior off-target profiles in these assessments, with minimal guide RNA-independent off-target activity and reduced Cas9-dependent off-target editing compared to base editors, attributable to the requirement for multiple independent hybridization events for successful editing [12] [9] [8].
Figure 2: Comprehensive experimental workflow for assessing bystander editing effects, from initial model selection through functional validation, as implemented in the rd12 mouse model study.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Research Reagents for Precision Editing Studies
| Reagent Category | Specific Examples | Function & Application |
|---|---|---|
| Base Editor Systems | ABEmax, ABE8e, ABE8eWQ, BE4max | Comparative assessment of editing precision across editor generations |
| Prime Editor Systems | PE2, PEmax, PE5, PE6 | Precision editing with minimal bystander effects |
| Delivery Vehicles | AAV2/9, Lipid Nanoparticles (LNPs), Dual AAV Split-Intein Systems | Efficient in vivo delivery of editing components |
| Cell Lines | HEK293T, K562 PEmax, PEmaxKO (MLH1-/-) | In vitro assessment of editing efficiency and specificity |
| Analysis Tools | CRISPResso2, Amplicon Sequencing, GUIDE-seq | Quantification of editing outcomes and off-target detection |
| Animal Models | rd12 mice (Rpe65 nonsense mutation) | In vivo functional validation of editing precision |
Emerging Solutions and Engineering Strategies
Recent advances in protein engineering have yielded novel approaches to enhance the precision of both base editing and prime editing systems, addressing the fundamental limitations of each platform.
Enhanced Specificity Base Editors
Directed evolution approaches have created context-specific base editors with narrowed editing windows to minimize bystander editing. A July 2025 study described the engineering of TadA deaminase variants through evolution of enzyme recognition "hotspots," generating cytosine-specific editors that function within defined nucleotide contexts. The resulting variants, including Tad-CBE4.1 with strong preference for ACG motifs, demonstrated significantly reduced bystander editing while maintaining on-target efficiency. These editors enable selective targeting of cytosines flanked by specific sequences, effectively mitigating the bystander effects that plague conventional base editors [35].
Advanced Prime Editing Systems
Prime editing efficiency has been substantially improved through multiple engineering strategies:
Engineered pegRNAs (epegRNAs): Incorporation of structured RNA motifs (evopreQ1, mpknot) at the 3' end of pegRNAs protects against degradation and improves editing efficiency 3-4-fold across multiple human cell lines [8] [19].
Mismatch Repair Inhibition: Co-expression of dominant-negative MLH1 (MLH1dn) in PE4 and PE5 systems suppresses mismatch repair, increasing editing efficiency from 20-40% to 50-80% in human cells [12] [19].
Dual Nicking Systems: PE3 and PE3b systems incorporate an additional sgRNA to nick the non-edited strand, encouraging the cellular repair machinery to use the edited strand as a template, increasing editing efficiency to 30-50% [12] [13].
Split Prime Editors: sPE systems separate nCas9 and RT components to overcome size limitations of viral delivery while maintaining editing precision [8].
These engineering advances have positioned prime editing as an increasingly viable platform for therapeutic applications requiring high precision, with recent optimized systems achieving >90% editing efficiency in benchmarked screens [19].
The comprehensive comparison of base editing and prime editing technologies reveals a fundamental trade-off between editing efficiency and precision. Base editors offer higher efficiency for specific transition mutations but suffer from significant bystander editing within their activity windows and deaminase-dependent off-target effects. In contrast, prime editors provide exceptional precision with minimal bystander editing and flexible editing capabilities but have historically faced efficiency challenges that are now being addressed through systematic engineering.
For therapeutic applications where precision is paramount - particularly for correcting point mutations without introducing collateral genetic changes - prime editing represents the superior platform despite its typically lower efficiency. The functional consequences of bystander editing demonstrated in the rd12 mouse model underscore the critical importance of precision in therapeutic genome editing. However, for research applications requiring efficient introduction of specific transition mutations or in contexts where delivery constraints favor smaller constructs, advanced base editors with narrowed editing windows may provide sufficient precision.
Future developments will likely further narrow the efficiency gap between these platforms while enhancing the precision of both systems. The optimal choice between base editing and prime editing ultimately depends on the specific application, target sequence context, delivery constraints, and precision requirements of each experimental or therapeutic scenario.
Optimizing pegRNA Design and Stability for Efficient Prime Editing
Prime editing represents a significant leap forward in precision genome editing, functioning as a versatile "search-and-replace" tool that can introduce targeted insertions, deletions, and all 12 possible base-to-base conversions without requiring double-strand DNA breaks (DSBs) or donor DNA templates [8] [3]. This technology centers on a prime editor (PE) complex—a fusion of a Cas9 nickase (nCas9) and a reverse transcriptase (RT)—guided by a specialized prime editing guide RNA (pegRNA) [8] [13]. The pegRNA not only directs the complex to the intended genomic locus but also encodes the desired genetic modification within its structure [13].
Despite its considerable promise, the transition of prime editing from a research tool to a reliable therapeutic modality has been hampered by challenges of variable and often low editing efficiency. A central bottleneck lies in the inherent instability and complex design requirements of the pegRNA molecule itself [8] [17]. Unlike conventional single-guide RNAs (sgRNAs), pegRNAs include extended 3' termini containing a primer binding site (PBS) and a reverse transcription template (RTT), making them more susceptible to cellular degradation and resulting in suboptimal performance [8] [13]. Consequently, optimizing pegRNA design and stability is not merely an incremental improvement but a fundamental prerequisite for achieving the high efficiency and reliability required for both basic research and clinical applications. Within the broader thesis comparing prime editing to base editing, a technology limited to four specific base transitions and prone to bystander edits, the resolution of pegRNA instability is what will ultimately unlock the full, unparalleled precision and versatility of prime editing [8] [3].
Engineering pegRNA Architecture for Enhanced Stability and Efficiency
The original pegRNA designs were prone to degradation by cellular exonucleases, which severely limited their effectiveness. To address this, researchers have developed engineered pegRNAs (epegRNAs) that incorporate structured RNA motifs at their 3' end. These motifs act as protective elements, shielding the pegRNA from degradation and thereby increasing its intracellular half-life and availability for the editing process [8].
Advanced pegRNA Modifications
- Structured RNA Motifs: Incorporating motifs such as evopreQ1 and mpknot at the 3' terminus of the pegRNA has been shown to improve prime editing efficiency by 3 to 4-fold across multiple human cell lines and primary fibroblasts. These structures prevent exonuclease-mediated degradation without increasing off-target effects [8].
- Alternative Stabilizing Motifs: Independent research efforts have yielded comparable stabilizing strategies. These include the use of a Zika virus exoribonuclease-resistant RNA motif (xr-pegRNA), the engineering of a G-quadruplex (G-PE), and the implementation of a stem-loop aptamer in a split prime editor (sPE) system [8].
- Circular RNA Templates: An innovative approach involves the use of circular prime editing template RNAs (petRNAs). This format inherently lacks exposed ends, conferring greater resistance to nucleases and helping to restore editing efficiency that can be lost due to degradation [32].
The following table summarizes the key pegRNA engineering strategies and their reported impacts on editing efficiency.
Table 1: pegRNA Engineering Strategies for Enhanced Stability and Efficiency
| Strategy | Key Feature | Reported Impact on Efficiency | Mechanism of Action |
|---|---|---|---|
| epegRNA [8] | EvopreQ1 or mpknot motif at 3' end | 3 to 4-fold increase | Protects 3' end from exonuclease degradation |
| xr-pegRNA [8] | Zika virus-derived resistant motif | Comparable improvement | Confers exonuclease resistance |
| G-PE [8] | G-quadruplex structure | Comparable improvement | Stabilizes pegRNA structure |
| petRNA [32] | Circular RNA template | Restores efficiency | Eliminates exposed ends susceptible to degradation |
| sPE [8] | Stem-loop aptamer | Comparable improvement | Enhances stability in split systems |
The proPE System: Decoupling Nicking and Templating Functions
A significant architectural innovation, termed proPE (prime editing with prolonged editing window), addresses limitations of the standard PE system by physically separating the nicking and templating functions onto two distinct RNA molecules [32].
- Essential Nicking Guide RNA (engRNA): A standard sgRNA that directs the prime editor to nick the target DNA site.
- Template Providing Guide RNA (tpgRNA): An RNA molecule containing the PBS and RTT, but with a truncated spacer sequence (11-15 nucleotides) that renders the associated Cas9 catalytically inactive. This tpgRNA binds nearby and provides the template efficiently [32].
This separation offers several key advantages that overcome specific bottlenecks of conventional PE, leading to an overall editing efficiency increase of up to 6.2-fold, particularly for edits that previously performed with low efficiency (<5%) [32]. The proPE system also broadens the effective editing window, bringing a larger fraction of pathogenic single nucleotide polymorphisms (SNPs) within reach [32].
The logical workflow and component relationships of the proPE system are illustrated below.
Diagram: The proPE system workflow, showing the separate roles of engRNA and tpgRNA.
Comparative Experimental Data on pegRNA Optimization
To objectively evaluate the performance of various optimization strategies, researchers systematically test editing efficiency across different genomic loci and modification types. The data below summarizes key findings from recent studies, providing a quantitative comparison of the strategies discussed.
Table 2: Comparative Editing Efficiencies of pegRNA Optimization Strategies
| Optimization Strategy | Editor Version | Target Locus / Cell Type | Edit Type | Reported Efficiency | Control (Standard PE) |
|---|---|---|---|---|---|
| epegRNA [8] | PE2 | Multiple human cell lines, primary fibroblasts | Point mutations | 3 to 4-fold increase | Baseline (1x) |
| proPE System [32] | proPE | Genomic targets in human cell lines | Edits with low PE efficiency | Up to 29.3% (from <5% baseline) | <5% |
| Systematic Delivery [18] | PEmax | HEK293T, HCT116, hPSCs | Multiple substitutions | Up to 80% (in cell lines), ~50% (in hPSCs) | Not specified |
| Stabilized pegRNA [18] | PEmax | Human pluripotent stem cells (naïve state) | Not specified | ~50% | Not specified |
The table demonstrates that integrated approaches, which combine improvements to the pegRNA with optimized editor proteins and delivery methods, can achieve exceptionally high editing efficiencies, even in challenging cell types like stem cells [18].
Detailed Experimental Protocols for pegRNA Assessment
For researchers to validate and build upon these findings, standardized protocols are essential. Below is a detailed methodology for a key experiment comparing the efficiency of standard pegRNAs versus epegRNAs.
Protocol: Evaluating epegRNA Performance
Objective: To quantitatively compare the editing efficiency of a standard pegRNA versus an epegRNA containing a 3' evopreQ1 motif at a specific genomic locus.
Materials:
- Plasmid DNA encoding the prime editor (e.g., pCMV-PE2 or pCMV-PEmax)
- Plasmid DNA for pegRNA expression (e.g., Lenti-TevopreQ1-Puro backbone)
- Oligonucleotides for cloning standard pegRNA and epegRNA sequences
- Human cell line (e.g., HEK293T)
- Transfection reagent
- Lysis buffer for genomic DNA extraction
- PCR primers for amplifying the target genomic locus
- Next-generation sequencing (NGS) library preparation kit
- NGS platform and data analysis software
Method:
- pegRNA Cloning: Clone the target spacer sequence, PBS, RTT, and the desired edit into the pegRNA expression plasmid. For the epegRNA, additionally clone the evopreQ1 sequence immediately following the RTT.
- Cell Transfection: Culture HEK293T cells and co-transfect them with a fixed amount of the prime editor plasmid and either the standard pegRNA or epegRNA plasmid. Include a negative control (e.g., transfected with editor plasmid only).
- Harvest and DNA Extraction: Incubate cells for 72 hours post-transfection to allow for editing. Harvest cells and extract genomic DNA using a standard protocol.
- Target Amplification and Sequencing: Amplify the target genomic region from the extracted DNA using PCR. Prepare the amplicons for NGS according to the manufacturer's instructions.
- Data Analysis: Sequence the libraries and analyze the results. Use bioinformatics tools to align sequences and calculate the percentage of reads containing the precise intended edit. Compare the editing efficiency between the standard pegRNA and epegRNA conditions.
Expected Outcome: The epegRNA condition is expected to show a statistically significant 3 to 4-fold increase in editing efficiency compared to the standard pegRNA, confirming the stabilizing effect of the 3' RNA motif [8].
The Scientist's Toolkit: Essential Research Reagents
Successful prime editing experiments, particularly those focused on pegRNA optimization, rely on a suite of specialized reagents and tools. The following table details key solutions for implementing the strategies discussed in this guide.
Table 3: Essential Research Reagents for pegRNA Optimization and Prime Editing
| Reagent / Solution | Function / Description | Example Use Case |
|---|---|---|
| Stabilized pegRNA Expression Vectors [18] [8] | Plasmid or viral backbones (e.g., Lenti-TevopreQ1-Puro) for expressing epegRNAs with 3' protective motifs. | Cloning and delivering epegRNAs for enhanced stability in difficult-to-transfect cells. |
| Optimized Prime Editor Plasmids [18] | Plasmids encoding advanced editors like PEmax, often with CAG or other strong promoters for high expression. | Provides the editor protein; used in co-transfection experiments to test novel pegRNA designs. |
| PiggyBac Transposon System [18] | A non-viral gene delivery system enabling stable genomic integration of the prime editor. | Generating isogenic cell lines with stable, sustained editor expression for high-throughput pegRNA screening. |
| Mismatch Repair Inhibitors [18] [13] | Co-expression of dominant-negative MLH1 (MLH1dn) to inhibit the mismatch repair pathway and prevent edit reversal. | Part of systems like PE5 to boost editing efficiency by modulating cellular repair responses. |
| Dual AAV Vector System [8] [32] | Adeno-associated virus vectors engineered to co-deliver large prime editing components, such as for the sPE or proPE systems. | Enables in vivo delivery of oversized prime editing machinery for therapeutic applications. |
| Lipid Nanoparticles (LNPs) [13] | Synthetic delivery particles for transient, efficient delivery of prime editing ribonucleoproteins (RNPs) or mRNA/pegRNA complexes. | Achieving high editing efficiency in vivo with reduced immunogenicity compared to viral vectors. |
The journey toward robust and predictable prime editing is inextricably linked to the optimization of the pegRNA. As this guide has detailed, strategic engineering of pegRNA stability through 3' motifs and innovative architectural changes like the proPE system can dramatically increase editing efficiency from single digits to over 80% in some contexts [18] [32]. The experimental data and protocols provided offer a roadmap for researchers to objectively compare and implement these strategies. As the field progresses, the integration of these optimized pegRNA designs with improved editor proteins, advanced delivery systems, and careful modulation of cellular repair pathways will be crucial to fully realizing the therapeutic potential of prime editing and solidifying its advantage in the precision genome editing landscape.
Precision genome editing technologies, particularly base editing and prime editing, have emerged as powerful tools for correcting pathogenic mutations without relying on double-strand DNA breaks (DSBs). Their therapeutic potential, however, is significantly modulated by cellular DNA repair pathways, with the mismatch repair (MMR) system playing a decisive role. MMR's primary function is to correct mispaired bases and small insertion-deletion loops (indels) that occur during DNA replication, but it also recognizes and processes the heteroduplex DNA intermediates generated during genome editing [36]. This review compares how MMR distinctly influences the efficiency and outcome fidelity of base editing and prime editing, synthesizing recent findings that reveal MMR as a major cellular hurdle that can be strategically overcome to optimize editing precision.
Primer on Mismatch Repair (MMR) Pathway Mechanics
The canonical MMR pathway is a highly conserved system for safeguarding genomic integrity. Its core components operate with specialized roles: the MutSα (MSH2-MSH6) complex primarily recognizes base-base mismatches and small (1-2 nucleotide) indels, while MutSβ (MSH2-MSH3) handles larger insertion-deletion loops [36] [37]. Following lesion recognition, MutLα (MLH1-PMS2) is recruited and, in a process requiring RFC and PCNA, introduces strand-specific nicks near the mismatch. These nicks serve as entry points for the EXO1 exonuclease, which excises the error-containing DNA segment. The resulting gap is then filled by DNA polymerase δ and sealed by DNA ligase I [36] [37]. This sophisticated repair machinery, when activated during genome editing, can profoundly alter the intended genetic outcome.
MMR Interplay with Base Editing Systems
Base Editor Architecture and Mechanism
Base editors are fusion proteins that typically combine a catalytically impaired Cas nuclease (dCas9 or nCas9) with a nucleotide deaminase enzyme. Cytosine Base Editors (CBEs) convert cytosine (C) to thymine (T) using a cytidine deaminase such as APOBEC1, while Adenine Base Editors (ABEs) convert adenine (A) to guanine (G) using an engineered tRNA adenosine deaminase (TadA) [38] [9]. The editing process creates a DNA heteroduplex containing a U•G (for CBEs) or I•T (for ABEs) mismatch, which the cellular MMR system must then resolve.
The following table summarizes the core components and MMR interactions of both editor types:
Table 1: Base Editing Systems and MMR Interplay
| Editor Type | Core Components | DNA Heteroduplex Created | MMR Recognition Complex | Typical MMR Outcome |
|---|---|---|---|---|
| CBE | nCas9 + Cytidine Deaminase (APOBEC1) + UGI [38] | U•G mismatch | MutSα (MSH2-MSH6) [36] | Resolution favors edited strand with UGI present [38] |
| ABE | nCas9 + Engineered Adenine Deaminase (TadA) [38] | I•T mismatch (I read as G) | MutSα (MSH2-MSH6) [36] | Resolution often favors edited strand [38] |
A critical optimization in CBE design is the inclusion of uracil DNA glycosylase inhibitor (UGI), which prevents base excision repair from reversing the U•G intermediate back to C•G, thereby increasing editing efficiency [38]. The U•G or I•T mismatches are recognized by the MutSα (MSH2-MSH6) complex [36]. While MMR can sometimes undesirably reverse base edits, the process generally exhibits higher efficiency and fewer indel byproducts compared to DSB-dependent methods [9].
MMR Interplay with Prime Editing Systems
Prime Editor Architecture and Mechanism
Prime editors consist of a Cas9 nickase (H840A) fused to an engineered reverse transcriptase (RT), programmed by a prime editing guide RNA (pegRNA) that specifies both the target site and encodes the desired edit [8] [13] [39]. The mechanism involves nicking the target DNA, followed by reverse transcription of the pegRNA template containing the edit, and finally resolution of the resulting DNA heteroduplex where one strand contains the original sequence and the other contains the new edit [39].
MMR as a Major Barrier to Prime Editing Efficiency
Unlike its more cooperative role in base editing, MMR acts as a potent inhibitor of prime editing efficiency. A comprehensive genetic screen targeting 32 DNA repair factors revealed that ablation of key MMR genes (MLH1, PMS2, MSH2, MSH3, EXO1) enhanced PE efficiency by 2- to 17-fold across various human cell lines, genomic loci, and edit types [37]. The accumulation of MMR proteins MLH1 and MSH2 at prime editing sites provides direct evidence of MMR's involvement in counteracting edit installation [37].
The following table quantifies the impact of MMR deficiency on prime editing efficiency across different experimental systems:
Table 2: Experimental Evidence of MMR Inhibition of Prime Editing
| Cell Line | MMR Deficiency | Edit Type | Fold Increase in PE Efficiency | Reference |
|---|---|---|---|---|
| HAP1 | MLH1 knockout | 5 bp deletion | 6.8× | [37] |
| HCT116 | MLH1/MSH3 deficient | 5 bp deletion | 2.8× | [37] |
| HEC59 | MSH2 deficient | 5 bp deletion | 6.6× | [37] |
| HEK293 | MLH1 knockout | 1 bp insertion | 14× | [37] |
| HEK293 | MLH1 knockout | 3 bp deletion | 7.5× | [37] |
The heteroduplex DNA formed during prime editing contains mismatches that are recognized as MMR substrates. The MMR system then preferentially excises the edited strand, viewing it as the "erroneous" one, thereby reversing the edit and reducing overall efficiency [37] [39]. This inhibitory effect is edit-type dependent, with larger indels being recognized by MutSβ (MSH2-MSH3) and smaller edits by MutSα (MSH2-MSH6) [37].
Strategic Engineering to Overcome MMR Inhibition
MMR Inhibition Strategies in Advanced Prime Editors
The understanding of MMR as a barrier to prime editing has driven the development of sophisticated engineering solutions. The most effective approach involves temporary inhibition of the MMR pathway during editing:
Table 3: Engineering Solutions to Overcome MMR Inhibition
| System | Engineering Strategy | Mechanism | Efficiency Improvement |
|---|---|---|---|
| PE4/PE5 | Co-expression of dominant-negative MLH1 (MLH1dn) [39] | Temporary inhibition of MutLα complex | 7.7× (PE4 vs PE2); 2.0× (PE5 vs PE3) [39] |
| epegRNA | 3' RNA pseudoknot structures [8] [39] | Protection of pegRNA from degradation; improves editing efficiency | 3-4× efficiency improvement [8] |
| PEmax | Codon optimization, additional NLS, Cas9 mutations [39] | Enhanced nuclear localization and nuclease activity | Variable but significant across targets [39] |
These engineered systems demonstrate that transient MMR inhibition, rather than permanent ablation, sufficiently enhances prime editing efficiency while maintaining genomic stability. The PE4 and PE5 systems specifically utilize a dominant-negative MLH1 mutant to temporarily disrupt the MutLα complex, providing a window of opportunity for the prime edit to be permanently incorporated without complete elimination of MMR's crucial genome maintenance functions [39].
Experimental Approaches for Studying MMR-Editor Interactions
Key Methodologies and Reagents
Investigating MMR's impact on editing technologies requires specialized experimental approaches:
Genetic Screens in Isogenic Cell Lines: The foundational evidence came from a targeted screen of 32 DNA repair genes in HAP1 cells, comparing PE efficiency in MMR-proficient and deficient backgrounds [37]. This approach enables systematic evaluation of how specific DNA repair pathways modulate editing outcomes.
MMR Inhibition via Dominant-Negative Mutants: Transient expression of dominant-negative MLH1 (MLH1dn) in the PE4/PE5 systems provides a controlled method to assess MMR impact without creating permanent MMR-deficient cell lines [39].
Amplicon Sequencing for Editing Quantification: Deep sequencing of edited genomic regions allows precise measurement of editing efficiency, unwanted indel rates, and editing fidelity [37].
Table 4: Essential Research Reagents for MMR-Editing Studies
| Reagent/Cell Line | Function/Application | Key Features |
|---|---|---|
| HAP1 MMR KO series | Isogenic background for genetic screens [37] | Near-haploid; enables clear functional assessment |
| HCT116 + chr3/5 | MMR-proficient control for MLH1-deficient HCT116 [37] | Chromosome complementation restores MMR |
| PE2, PE3 systems | Base prime editing platforms [39] | Enable comparison of editing with/without strand nicking |
| PE4, PE5 systems | MMR-optimized prime editors [39] | Incorporate MLH1dn for enhanced efficiency |
| pegRNA/epegRNA | Prime editing guide RNAs [8] [39] | epegRNAs include stabilizing 3' structures |
The distinct interplay between MMR and precision genome editing technologies reveals a critical determinant of editing success. While base editing benefits from MMR activity to resolve intermediate mismatches, prime editing faces significant inhibition from this pathway. Strategic engineering, particularly through temporary MMR suppression in advanced prime editor systems, has dramatically improved editing efficiencies. These insights underscore that understanding and overcoming cellular DNA repair mechanisms is not merely an optimization step but a fundamental requirement for realizing the full therapeutic potential of precision genome editing. As these technologies advance toward clinical application, managing the MMR response will remain crucial for achieving predictable, efficient, and safe editing outcomes across diverse cell types and genetic contexts.
The development of prime editing represents a paradigm shift in genome engineering, moving beyond the limitations of earlier technologies that rely on double-strand breaks (DSBs). Unlike traditional CRISPR-Cas9 systems that induce DSBs and rely on error-prone repair mechanisms, prime editors enable precise "search-and-replace" genome editing without requiring donor DNA templates or creating double-strand breaks [8] [3]. This technology has rapidly evolved through several generations—from the foundational PE2 to the optimized PEmax and the newly developed vPE—each offering significant improvements in editing efficiency and precision while minimizing unwanted byproducts [8] [40].
The broader context of precision genome editing research involves a critical comparison between base editing and prime editing technologies. While base editors can efficiently perform single-nucleotide conversions (C-to-T or A-to-G), they are limited to four of the twelve possible base-to-base transitions and can suffer from bystander edits within their activity window [41] [3]. Prime editing, in contrast, theoretically enables all 12 possible base substitutions, along with small insertions and deletions, offering substantially greater versatility [8] [13]. This comparison framework is essential for understanding the engineering progress from PE2 to vPE, as each advancement addresses specific limitations in the precision-editing landscape.
Architectural Foundations of Prime Editing Systems
Core Components and Mechanism
Prime editing systems consist of several essential components that work in concert to enable precise genome modifications. The core architecture includes a Cas9 nickase (H840A) fused to an engineered reverse transcriptase (RT) from Moloney Murine Leukemia Virus (MMLV), programmed with a specialized prime editing guide RNA (pegRNA) [8] [13]. The pegRNA serves dual functions: it directs the Cas9 nickase to the target genomic locus and also encodes the desired edit through its reverse transcription template (RTT) and primer binding site (PBS) regions [13].
The editing mechanism occurs through a sophisticated multi-step process: (1) The PE complex binds to the target DNA, and the Cas9 nickase creates a single-strand nick in the non-target DNA strand; (2) The released 3' hydroxyl group primes reverse transcription using the pegRNA's RTT sequence; (3) The newly synthesized edited strand must then compete with the original DNA flap for incorporation into the genome through cellular repair mechanisms [8] [40]. This final strand competition step represents a critical point where editing efficiency can be compromised and unwanted indel errors may occur [40].
The Critical Role of pegRNA Engineering
The pegRNA is a complex molecule typically 120-145 nucleotides in length, significantly longer than standard CRISPR guide RNAs [13]. This extended length presents challenges for synthesis, delivery, and stability. Initial prime editing systems suffered from pegRNA degradation, which limited editing efficiency. This limitation prompted the development of engineered pegRNAs (epegRNAs) that incorporate structured RNA motifs such as evopreQ, mpknot, or exoribonuclease-resistant motifs at their 3' ends to protect against degradation [8]. These stabilizing modifications improve prime editing efficiency by 3-4-fold across multiple human cell lines and primary human fibroblasts without increasing off-target effects [8].
More recent advances include La-accessible pegRNAs, which incorporate polyU tails that enhance interaction with the RNA-binding protein La, significantly boosting editing efficiency when used with the PE7 system [42]. In zebrafish models, this combination achieved editing efficiencies of up to 15.99%—a 6.81- to 11.46-fold improvement over earlier PE2 systems [42].
Evolution of Prime Editing Systems: From PE2 to vPE
PE2: The Foundation for Optimization
The development of PE2 followed the initial PE1 system through optimization of the reverse transcriptase component fused to the Cas9 nickase (H840A) [8]. These modifications enhanced the thermostability, processivity, and affinity for RNA-DNA hybrid substrates, resulting in improved editing outcomes across a broader range of genomic contexts [8]. While PE2 demonstrated the feasibility of precise genome editing without double-strand breaks, its efficiency remained context-dependent and limited in some applications, prompting further engineering efforts.
PEmax: Enhanced Efficiency and Versatility
Building upon PE2, the PEmax system incorporated additional mutations to optimize editor performance, though the specific mutations are not detailed in the available literature [40]. PEmax represents a significant step forward in prime editing capability, with enhanced editing efficiency across diverse genomic loci. However, despite these improvements, PEmax systems still generated unwanted indel byproducts at frequencies that raised concerns for therapeutic applications [40]. These error rates, ranging from approximately one error in seven edits to one in 121 edits for different editing modes, highlighted the need for further refinement of the technology [43].
vPE: Dramatically Reduced Errors Through Nick Positioning
The most recent advancement in prime editing technology comes with the development of vPE (next-generation prime editor), which addresses the critical challenge of indel formation during the editing process [40] [43]. The innovation centers on engineering Cas9 nickase variants with relaxed nick positioning, specifically through mutations at residues R780, K810, K848, K855, R976, and H982 [40].
These mutations promote degradation of the competing 5' DNA strand (non-target strand), which biases the repair process toward incorporation of the edited 3' strand [40]. This strategic approach significantly reduces the formation of indel errors that previously limited prime editing applications. The vPE system demonstrates remarkable improvements in precision, with edit:indel ratios as high as 543:1 in high-precision editing modes—representing up to a 60-fold reduction in errors compared to previous editors [40] [43].
Table 1: Comparative Performance of Prime Editor Generations
| Editor Version | Key Innovations | Editing Efficiency | Error Rate (Indels) | Edit:Indel Ratio |
|---|---|---|---|---|
| PE2 | Optimized RT domain | Context-dependent, moderate | Significant | Limited data available |
| PEmax | Additional protein engineering | Improved over PE2 | 1 error per 7-121 edits [43] | Lower than vPE |
| vPE | Relaxed nick positioning (e.g., K848A-H982A) | Comparable to PEmax | 1 error per 101-543 edits [43] | Up to 543:1 [40] |
Experimental Protocols and Validation
Methodology for Evaluating Prime Editor Performance
The quantitative comparison between prime editor generations relies on standardized experimental protocols. Typical validation involves transfection of HEK293T cells with editor plasmids and corresponding pegRNAs, followed by next-generation sequencing of target loci to quantify editing efficiency and indel formation [40]. Key experimental steps include:
- Plasmid Construction: Prime editor constructs (PE, PEmax, pPE, vPE) are cloned into mammalian expression vectors with appropriate regulatory elements [40].
- Cell Transfection: HEK293T cells are transfected with editor plasmids and pegRNA vectors using standard methods (e.g., lipid-based transfection) [40].
- Target Selection: Editing is evaluated at multiple genomic loci (e.g., CXCR4, EMX1, TGFB1, KRAS) with diverse sequence contexts to assess generalizability [40].
- Sequencing Analysis: Genomic DNA is extracted 72-96 hours post-transfection, target sites are amplified by PCR, and products are sequenced using Illumina platforms [40].
- Data Processing: Editing efficiency is calculated as the percentage of reads containing the desired edit, while indel rates are quantified as the percentage of reads with insertions or deletions at the target site [40].
For the vPE system, additional assays measured nick shift frequency and flap degradation to correlate structural changes with functional improvements [40]. The flap degradation assay specifically involved creating paired nicks to produce homology deletions, where degraded nicked ends inhibit deletions, allowing quantification of end stability [40].
Ribonucleoprotein (RNP) Delivery in Model Systems
Recent advances have demonstrated the effectiveness of ribonucleoprotein (RNP) complex delivery for prime editing, particularly in challenging model systems like zebrafish. One protocol achieved 15.99% editing efficiency in zebrafish embryos using PE7 protein complexed with La-accessible pegRNAs [42]. The experimental workflow involved:
- RNP Complex Formation: Incubating PE7 protein with La-accessible pegRNAs to form RNP complexes [42].
- Microinjection: Delivering 2 nL of RNP complexes (750 ng/μL PE7 + 240 ng/μL pegRNA) into zebrafish embryos at the one-cell stage [42].
- Efficiency Quantification: Extracting genomic DNA at 2 days post-fertilization, followed by amplicon sequencing and analysis [42].
This approach achieved a 6.81- to 11.46-fold improvement over PE2 systems and successfully generated a visible phenotype (melanin reduction) through precise tyr P302L mutation, demonstrating the practical application of advanced prime editing systems [42].
Research Reagent Solutions for Prime Editing
Table 2: Essential Research Reagents for Prime Editing Experiments
| Reagent/Category | Specific Examples | Function and Application |
|---|---|---|
| Prime Editor Plasmids | PE2, PEmax, PE7, vPE expression vectors | Provide the engineered editor proteins for mammalian cell expression |
| pegRNA Components | Standard pegRNAs, epegRNAs, La-accessible pegRNAs (with polyU tails) | Direct targeting and encode desired edits; stabilized versions improve efficiency |
| Delivery Tools | Lipid nanoparticles (LNPs), electroporation, AAV vectors (for split systems) | Enable intracellular delivery of editing components |
| Nicking gRNAs | PE3, PE3b systems | Secondary gRNAs to nick non-edited strand and increase efficiency |
| MMR Inhibitors | MLH1dn (in PE5 system) | Block mismatch repair to prevent reversal of edits |
| Stabilizing Proteins | La protein (enhances PE7 activity) | RNA-binding proteins that improve pegRNA stability and editing efficiency |
| Analysis Tools | Barcoded PCR primers, NGS platforms | Quantify editing efficiency and indel formation |
Visualizing Prime Editing Evolution and Performance
The diagram below illustrates the evolutionary pathway from PE2 to vPE, highlighting key innovations and performance characteristics at each stage:
Diagram 1: Prime editing evolution from PE2 to vPE shows progressive error reduction.
The following diagram illustrates the critical mechanism of nick positioning relaxation that enables vPE's improved precision:
Diagram 2: vPE mechanism shows how relaxed nick positioning reduces errors.
The evolution from PE2 to vPE represents a remarkable trajectory of innovation in precision genome editing. Each generation has addressed specific limitations: PE2 established a functional foundation, PEmax enhanced efficiency, and vPE dramatically reduced error rates through sophisticated protein engineering that exploits DNA repair mechanisms [40] [43]. The progressive improvement in edit:indel ratios—reaching up to 543:1 with vPE—demonstrates the potential of prime editing for therapeutic applications where precision is paramount [40].
These engineering advances position prime editing as a versatile technology capable of addressing a broad spectrum of genetic variants. Computational models suggest prime editing could theoretically correct up to 89% of known pathogenic human genetic variants [3]. The first clinical trials of prime editing therapies are already underway, with FDA approval of an investigational new drug application for PM359, a prime editing treatment for chronic granulomatous disease [16]. This rapid translation from basic research to clinical application highlights the transformative potential of these continuing improvements in prime editing technology.
Future development will likely focus on further enhancing editing efficiency, expanding targetability, and addressing delivery challenges—particularly for in vivo applications. As the precision and versatility of prime editing continue to improve, this technology promises to unlock new therapeutic strategies for genetic disorders that have previously been intractable to conventional gene therapy approaches.
The advent of precision genome editing tools, namely base editing and prime editing, has heralded a new era in therapeutic development for genetic disorders. These technologies surpass traditional CRISPR-Cas9 systems by enabling precise nucleotide changes without inducing double-strand DNA breaks, thereby reducing unwanted mutagenic outcomes [3]. However, their clinical translation is critically hampered by a shared, central challenge: the large physical size of the editor complexes, which complicates packaging and delivery into target cells using conventional viral vectors [3] [8]. This article objectively compares the distinct delivery challenges and the evolving strategies for both base and prime editing platforms, providing a foundational resource for researchers and drug development professionals navigating this complex landscape.
Base editing and prime editing represent two distinct approaches to precision genome editing, each with unique architectures and delivery implications.
Base Editing: Base editors utilize a catalytically impaired Cas protein (a nickase or one completely devoid of cutting activity) fused to a deaminase enzyme. This complex catalyzes the direct chemical conversion of one base into another (e.g., C to T or A to G) without a double-strand break [3] [13]. While this avoids the genotoxic risks of traditional CRISPR, the fusion protein is substantially larger than the Cas9 protein alone.
Prime Editing: The prime editing system is more complex. It employs a Cas9 nickase fused to an engineered reverse transcriptase (RT), programmed by a specialized prime editing guide RNA (pegRNA) [8] [13]. The pegRNA is notably longer than a standard sgRNA because it must contain both the spacer sequence and an extended template encoding the desired edit. This multi-component system, particularly the large PE protein and the lengthy pegRNA, presents a significant delivery obstacle [3] [13].
The table below summarizes the core components and their relative contribution to the delivery challenge for each system.
Table 1: System Architecture and Size Comparison
| Feature | Base Editing System | Prime Editing System |
|---|---|---|
| Core Editor Protein | Cas nickase/deadCas + Deaminase | Cas9 nickase (H840A) + Reverse Transcriptase [8] [13] |
| Guide RNA | Standard sgRNA (shorter) | Prime Editing Guide RNA (pegRNA) - ~120-145+ nucleotides [13] |
| Primary Delivery Hurdle | Large fusion protein exceeds AAV cargo capacity [7]. | Very large fusion protein and a long, structured pegRNA complicate co-delivery [3] [8]. |
Quantitative Analysis of Delivery Strategies and Efficiencies
Developing effective delivery strategies is paramount for both research and therapeutic applications. The following table synthesizes key approaches, their methodologies, and performance data as evidenced in recent literature.
Table 2: Comparison of Delivery Strategies and Experimental Outcomes
| Delivery Strategy | Experimental Protocol & Methodology | Reported Efficiency/Outcome | Key Advantages & Limitations |
|---|---|---|---|
| Dual AAV Vectors (Split-intein) | Protocol: The editor protein is split into two halves, each packaged into a separate AAV vector, with split intein domains that reconstitute the full protein post-delivery. Often used for both base and prime editors [8] [7]. | Base Editing: Successfully used in mouse models; editing efficiencies vary widely by target and tissue [7].Prime Editing: Enables in vivo editing in mouse liver; efficiencies can be lower than base editing due to complex reconstitution [8]. | Adv.: Bypasses AAV cargo limit.Disadv.: Reconstitution inefficient; potential for immune response. |
| Virus-Like Particles (VLPs) & Non-Viral Vectors | Protocol: Engineered VLPs or lipid nanoparticles (LNPs) are used to package editor mRNA or ribonucleoprotein (RNP) complexes. This is a rapid, transient delivery method [13] [44]. | LNP delivery of base editor mRNA is an emerging strategy noted for its therapeutic potential [7]. MIT's vPE system demonstrated reduced errors in mouse and human cells via non-viral delivery [44] [15]. | Adv.: High transient expression; suitable for therapeutics.Disadv.: Can have lower efficiency than viral vectors in some tissues. |
| Stable Cell Line Expression | Protocol: Creating clonal cell lines (e.g., K562, HEK293) that constitutively express the editor protein (e.g., PE2, PEmax). Editing is then initiated by delivering only the pegRNA [19]. | A benchmarked study showed precise prime editing efficiencies reaching ~95% in an MMR-deficient (PEmaxKO) cell line over 28 days [19]. | Adv.: High editing yield; ideal for ex-vivo and screening applications.Disadv.: Not suitable for in vivo therapy; risk of chronic off-target activity. |
| Engineered Compact Editors | Protocol: Using smaller Cas orthologs (e.g., SaCas9) or splitting the editor into smaller parts to facilitate packaging. The "sPE" system allows nCas9 and RT to function independently [8] [12]. | The sPE system successfully edited the mouse liver β-catenin gene and corrected a mutation in a model of type I tyrosinemia using a dual AAV system [8]. | Adv.: Expands delivery options, especially for AAVs.Disadv.: May have reduced editing scope or efficiency. |
Visualizing Strategic Approaches and Experimental Workflows
The following diagrams map the logical relationships between delivery challenges and strategies, as well as a key experimental workflow from recent research.
Strategic Framework for Delivery Challenges
Workflow: High-Efficiency Prime Editing in Stable Cell Lines
The Scientist's Toolkit: Key Reagents and Research Solutions
Successful experimentation in this field relies on a suite of specialized reagents and tools. The table below details essential materials for implementing advanced delivery and editing protocols.
Table 3: Essential Research Reagents for Advanced Editing Studies
| Reagent / Tool Name | Function & Application in Research |
|---|---|
| epegRNA (e.g., tevopreQ1 motif) | Engineered pegRNA with a 3' RNA motif that enhances stability and increases prime editing efficiency by 3-4-fold by protecting against exonuclease degradation [19] [8]. |
| PEmax Protein | An optimized version of the PE2 prime editor protein with improved nuclear localization and expression, leading to higher editing efficiencies across multiple targets [19]. |
| MLH1dn (Dominant-negative MLH1) | A key reagent for inhibiting the DNA mismatch repair (MMR) pathway. Co-expression with prime editors (as in PE4/PE5 systems) significantly boosts editing efficiency by preventing the repair machinery from rejecting the newly edited strand [19] [12]. |
| Split-intein AAV System | A delivery solution where the large editor protein is divided into two parts, each fused to split intein domains and packaged into separate AAVs. Reconstitution in the host cell produces the full-length, functional protein, overcoming AAV packaging constraints [8] [7]. |
| sPE (Split Prime Editor) System | A prime editor architecture where the nCas9 and reverse transcriptase components are delivered separately. This simplifies packaging into dual AAV vectors and has been shown to be effective for in vivo editing in mouse models [8]. |
The delivery of large editor complexes remains a formidable but not insurmountable barrier in the development of precision gene therapies. Base editors and prime editors, while sharing the challenge of size, are being advanced through a combination of parallel and distinct strategies. Dual-vector AAV systems and the development of compact, split editors are direct responses to the physical packaging problem. Meanwhile, non-viral delivery and the use of stable cell lines offer powerful alternatives for both transient ex vivo applications and high-throughput research. The choice of strategy is highly context-dependent, influenced by the target tissue, the specific editor, and the ultimate therapeutic goal. As the field progresses, the continued refinement of these delivery strategies will be just as critical as the development of the editors themselves for unlocking the full clinical potential of precision genome editing.
Benchmarking Efficiency, Precision, and Fidelity
In the rapidly advancing field of precision genome editing, two technologies have emerged as front-runners: base editing and prime editing. Both promise to correct genetic mutations without inducing double-strand DNA breaks, a significant limitation of earlier CRISPR-Cas9 nuclease approaches. However, accurately quantifying the efficiency and purity of the edits they produce is paramount for comparing their performance and guiding therapeutic development. This guide examines the critical role of amplicon sequencing as the benchmark method for the precise, quantitative analysis of base and prime editing outcomes, providing researchers with the data and methodologies needed for robust evaluation.
The Imperative for Precision in Genome Editing
The evolution from nuclease-based gene editing to more precise, "next-generation" techniques represents a paradigm shift in our ability to manipulate the genome.
- The Double-Strand Break Problem: Traditional CRISPR-Cas9 nucleases act by creating double-strand breaks (DSBs) in DNA. The cell's repair of these breaks often results in a spectrum of insertion and deletion mutations (indels). While useful for gene disruption, this stochastic mixture of outcomes is a major drawback for precision applications, as the sequence of indels cannot be specified and DSBs can trigger undesirable cellular responses, including chromosomal rearrangements and activation of p53 [25].
- The Rise of Precision Editors:
- Base Editors (BEs) use a catalytically impaired Cas protein fused to a deaminase enzyme to directly convert one base pair to another—specifically, cytosine to thymine (C•G to T•A) or adenine to guanine (A•T to G•C)—without making a DSB. While highly efficient, their application is confined to these specific transition mutations and they can cause unwanted "bystander" edits to adjacent bases within a small activity window [8] [25] [2].
- Prime Editors (PEs) offer a more versatile solution. A prime editor is a fusion of a Cas9 nickase and a reverse transcriptase, programmed by a specialized prime editing guide RNA (pegRNA). This system can mediate all 12 possible base-to-base conversions, as well as small targeted insertions and deletions, all without requiring DSBs or donor DNA templates. This grants prime editing exceptionally high versatility and editing purity compared to both nucleases and base editors [8] [13] [25].
The very nature of these precise tools—designed to create specific, single-nucleotide or small-sequence changes amidst a vast background of unedited genomic DNA—demands a quantification method of commensurate precision. This is where amplicon sequencing becomes indispensable.
Amplicon Sequencing: The Gold Standard for Quantification
Amplicon sequencing, a targeted approach where specific genomic regions of interest are amplified via PCR and subjected to high-throughput sequencing, has established itself as the gold standard for evaluating genome editing outcomes due to its sensitivity, quantitative nature, and scalability [45] [46].
The following diagram illustrates the typical workflow for using amplicon sequencing to quantify gene editing outcomes, from sample preparation to data analysis.
Key Advantages for Editing Analysis
- High Sensitivity and Accuracy: Amplicon sequencing can detect editing events present at very low frequencies (well below 1%) within a heterogeneous cell population, providing a true measure of editing efficiency [46].
- Comprehensive Outcome Characterization: It reveals the complete spectrum of DNA changes at the target site. This includes not only the desired edit but also precise byproducts such as:
- Reliance on Reference Materials: To ensure accuracy, the Genome in a Bottle (GIAB) reference materials from the National Institute of Standards and Technology (NIST) provide high-confidence benchmark datasets. These allow labs to validate their amplicon sequencing assays and bioinformatics pipelines by comparing their results against a known "truth set," thereby generating standardized performance metrics like sensitivity and precision [45].
Quantifying Base Editing Outcomes
Base editing efficiency is influenced by the guide RNA (gRNA) sequence and the local sequence context of the target nucleotide. Amplicon sequencing is critical for measuring both the primary edit and unwanted bystander effects.
Experimental Protocol for Base Editing Analysis
- Design: Design PCR primers that flank the target base editing window, typically allowing for a 30-50 bp margin on either side.
- Amplification: Amplify the target region from purified genomic DNA. Using a high-fidelity polymerase is crucial to minimize PCR-introduced errors.
- Library Preparation & Sequencing: Prepare sequencing libraries from the amplified products (amplicons) using a platform like Illumina. The high coverage depth (often >1000x) enables accurate frequency calculations for each observed variant [21].
- Analysis: Use computational tools to align sequences to the reference genome and call variants. The base editing efficiency is calculated as the percentage of sequencing reads containing the intended base conversion out of all reads covering that position.
Characterizing Base Editing with Deep Learning
Large-scale amplicon sequencing data has enabled the development of sophisticated predictive models. For instance, the CRISPRon-BE tool is a deep learning model trained on amplicon sequencing data from over 11,000 gRNAs for both Adenine Base Editors (ABEs) and Cytosine Base Editors (CBEs). This model simultaneously predicts gRNA efficiency and the frequency of all possible editing outcomes within the activity window, demonstrating how amplicon data is fundamental to advancing the field [21].
The table below summarizes key quantitative findings from base editing studies analyzed via amplicon sequencing.
Table 1: Quantitative Analysis of Base Editing Outcomes via Amplicon Sequencing
| Base Editor Type | Core Editing Window | Primary Conversion | Bystander Edit Frequency | Indel Frequency | Key Insight |
|---|---|---|---|---|---|
| ABE7.10 (Adenine) | Positions 4-8 | A•T to G•C (~97% of edits) | Low | ~2.1% | Highly precise with minimal byproducts [21]. |
| BE4 (Cytosine) | Positions 4-8 | C•G to T•A (~92% of edits) | Moderate (C•G to G•C ~3.5%) | ~3.0% | Less strict than ABEs, with more frequent bystander transversions [21]. |
Quantifying Prime Editing Outcomes
Quantifying prime editing is complex due to the diversity of possible edits and the potential for byproducts like indels and incomplete edits. Amplicon sequencing is the only method capable of providing a full picture of this heterogeneity.
Experimental Protocol for Prime Editing Analysis
The foundational protocol is similar to that for base editing, but special attention must be paid to the primer design to ensure the primers bind outside the homologous regions used in the pegRNA's template to avoid amplification bias. The analysis pipeline must be tailored to detect the specific DNA sequence change encoded by the pegRNA's reverse transcriptase template (RTT).
Case Study: Enhancing Prime Editing with proPE
A 2025 study, "ProPE expands the prime editing window and enhances gene editing efficiency," leveraged amplicon sequencing (Amplicon deep sequencing of human cell lines) to rigorously quantify the improvements offered by a new system called proPE. proPE uses a second, non-cleaving sgRNA to deliver the editing template, which addresses several bottlenecks of traditional PE [32].
The study demonstrated that for edits where traditional PE was inefficient (editing efficiency <5%), the proPE system resulted in an overall 6.2-fold increase in editing efficiency, achieving up to 29.3% efficiency for these challenging targets. This data, quantifiable only through deep amplicon sequencing, was pivotal in validating the new system's performance [32].
Table 2: Quantitative Comparison of Prime Editing Systems via Amplicon Sequencing
| Editing System | Typical Edit Types | Reported Efficiency Range | Key Byproducts | Notable Advantage |
|---|---|---|---|---|
| PE2 | Point mutations, small insertions/deletions | Variable, can be low (<5-10%) [32] [25] | Indels, incomplete edits | Simplicity; improved product purity over HDR [25]. |
| PE3/PE3b | Point mutations, small insertions/deletions | Up to ~2x PE2 efficiency [8] [25] | Indels (can be higher than PE2) | Increased editing efficiency via nicking of non-edited strand [8] [25]. |
| proPE (2025) | Point mutations, edits beyond typical window | 6.2-fold increase for low-efficiency PE targets (up to 29.3%) [32] | Reduced re-nicking and re-binding [32] | Less optimization needed; broader editing window; higher specificity [32]. |
Successful quantification of editing outcomes relies on a suite of key reagents and reference materials.
Table 3: Essential Research Reagent Solutions for Quantifying Editing Outcomes
| Item / Solution | Function / Explanation | Example Use Case |
|---|---|---|
| High-Fidelity DNA Polymerase | Amplifies the target genomic locus with minimal errors, preventing false positives in sequencing data. | Critical for all amplicon sequencing workflows to ensure data fidelity [47]. |
| Illumina Amplicon Library Prep Kits | Streamlined kits (e.g., Illumina Microbial Amplicon Prep - iMAP) for preparing sequencing libraries from PCR amplicons. | Used in optimized workflows for high-throughput pathogen surveillance, adaptable for editing analysis [47]. |
| NIST Genome in a Bottle (GIAB) Reference Materials | Provides benchmark genomic DNA and high-confidence variant calls to validate assay accuracy and bioinformatics pipelines. | Used to calculate performance metrics like sensitivity and precision for a targeted sequencing panel [45]. |
| Structured pegRNAs (epegRNAs) | pegRNAs with added RNA motifs (e.g., evopreQ) at the 3' end to resist degradation, boosting prime editing efficiency. | Can increase prime editing efficiency 3-4-fold, as measured by amplicon sequencing [8]. |
| Prime Editor Protein Variants | Engineered editors like PE2 (with optimized RT) and PE5 (with MMR inhibition) for improved efficiency and product purity. | PE2 offers 1.6-5.1x higher efficiency than PE1; PE5 reduces mismatch repair reversal of edits [13] [25]. |
The following diagram outlines the core computational steps for analyzing amplicon sequencing data to generate the final quantitative metrics that researchers rely on.
In the direct comparison between base editing and prime editing, amplicon sequencing provides the critical data needed to weigh their respective strengths and limitations. Base editors offer remarkable efficiency and purity for their specific, narrow set of transition mutations. In contrast, prime editing stands out for its exceptional versatility, capable of installing a much broader range of precise changes, albeit often with more variable efficiency that next-generation systems like proPE are seeking to address.
Across all these technologies, amplicon sequencing remains the unassailable gold standard for quantification. Its unparalleled sensitivity, capacity for comprehensive outcome characterization, and role in validating predictive models and new editor systems make it an indispensable tool. For researchers and drug developers aiming to bring precision gene editing into the therapeutic realm, robust amplicon sequencing protocols are not just best practice—they are a fundamental requirement for generating the conclusive data that will propel the field forward.
The journey from traditional CRISPR-Cas9 systems to more precise gene editing technologies has been largely driven by the need to address the "indel problem"—the unwanted insertions and deletions that frequently arise from DNA double-strand breaks (DSBs). These indels represent a significant safety concern for therapeutic applications, as they can lead to unpredictable mutagenic outcomes, including genomic instability and potential oncogenic events [8]. While initial CRISPR systems revolutionized biotechnology by enabling targeted genome modification, their reliance on inducing DSBs and subsequent repair through error-prone non-homologous end joining (NHEJ) pathways results in a high frequency of these undesirable byproducts [3] [8].
Base editing and prime editing emerged as revolutionary technologies designed to minimize or eliminate DSBs, thereby offering a potential solution to the indel problem. Base editing, developed by Dr. David Liu's team, utilizes a modified Cas9 enzyme that cuts only one DNA strand or none at all, fused to a deaminase enzyme that directly converts one base to another without creating DSBs [3] [13]. Prime editing, a more recent innovation, functions as a "search-and-replace" tool that uses a Cas9 nickase fused to a reverse transcriptase, guided by a specialized prime editing guide RNA (pegRNA) to directly write new genetic information into a target DNA site [3] [8]. Both technologies represent significant advancements over traditional CRISPR-Cas9, but they differ substantially in their mechanisms, capabilities, and particularly in their byproduct profiles, including their propensity to generate indels.
Mechanisms of Byproduct Generation: Fundamental Differences
Base Editing Mechanisms and Bystander Effects
Base editors operate through a sophisticated mechanism that combines targeted DNA binding with enzymatic base conversion. Cytosine base editors (CBEs) convert cytosine (C) to thymine (T), while adenine base editors (ABEs) convert adenine (A) to guanine (G) [13]. The process begins with the base editor complex binding to a specific DNA sequence guided by a single guide RNA (sgRNA). Unlike traditional CRISPR-Cas9, the Cas9 component in base editors is modified to form a nickase (nCas9) that cuts only one DNA strand or is catalytically impaired (dCas9) to make no cuts at all [3]. The fused deaminase enzyme then acts on a specific base within a defined "editing window" typically 4-5 nucleotides in length, converting it to another base through deamination [8].
This mechanism successfully avoids DSBs but introduces its own unique byproducts. The most significant issue is bystander editing, where the base editor makes unwanted changes to neighboring bases within the editing window [3] [8]. For example, when targeting a specific cytosine residue, other cytosines within the editing window may also be deaminated, leading to multiple, potentially undesirable base changes alongside the intended edit. Although base editing significantly reduces indel formation compared to traditional CRISPR-Cas9, indels still occur at low frequencies, particularly because the nickase activity can still trigger some degree of error-prone repair [48]. Additionally, base editors can exhibit off-target mutations in both DNA and RNA due to the deaminase activity of APOBEC and TadA enzymes used in these editors [8].
Prime Editing and Its Refined Approach
Prime editing represents a more versatile approach that further minimizes undesirable byproducts. The core prime editing system consists of a Cas9 nickase (H840A) fused to an engineered reverse transcriptase (RT), programmed with a specialized pegRNA [8] [12]. The pegRNA not only specifies the target site but also contains a reverse transcription template (RTT) encoding the desired edit. The editing process begins when the prime editor complex binds to the target DNA and the nCas9 nicks the non-target strand. The exposed 3'-hydroxyl group acts as a primer for the RT to synthesize new DNA using the RTT of the pegRNA as a template [8]. This creates a branched intermediate structure where the edited and unedited strands coexist, which is subsequently resolved by cellular repair mechanisms that favor the incorporation of the edited strand.
The prime editing mechanism inherently produces fewer byproducts than both traditional CRISPR and base editing. Since it doesn't create DSBs and doesn't rely on deaminase enzymes, it avoids both the indel problem associated with DSB repair and the bystander editing characteristic of base editors [8] [12]. However, prime editing is not completely free of byproducts. The initial versions of prime editors could still generate low levels of indels, primarily due to the potential for the nCas9 to inadvertently generate DSBs or from imperfect resolution of the DNA flap intermediates during the editing process [8]. Later iterations of prime editors have incorporated additional mutations to further minimize these undesirable byproducts.
Table 1: Fundamental Mechanisms and Primary Byproducts of Gene Editing Technologies
| Technology | Core Mechanism | DNA Cleavage Type | Primary Byproducts | Key Limitations |
|---|---|---|---|---|
| CRISPR-Cas9 | Cas9 nuclease creates DSBs, repaired by NHEJ or HDR | Double-strand breaks | High indels, large deletions, chromosomal rearrangements | Unpredictable outcomes, high genotoxic risk |
| Base Editing | Deaminase fused to nCas9 or dCas9 directly converts bases | Single-strand nick or no cut | Bystander edits, low-frequency indels, RNA off-targets | Restricted to 4 of 12 possible base transitions |
| Prime Editing | nCas9-Reverse Transcriptase fusion uses pegRNA template | Single-strand nick | Very low indels, imperfect edit incorporation | Large size complicating delivery, variable efficiency |
Quantitative Comparison of Byproduct Profiles
Direct Comparative Studies and Efficiency Metrics
Recent advances in both base editing and prime editing have yielded improved versions with enhanced precision, enabling a more direct comparison of their byproduct profiles. For base editing, the development of QBEmax—a sequence-permuted and internally protected base editor—represents a significant step forward in minimizing undesirable byproducts. In comparative studies, QBEmax demonstrated dramatically reduced indel formation, with average indel frequencies of just 1.2% ± 0.2% across 17 genomic sites, representing a 56.5% reduction compared to previous editors like BE4max [48]. Furthermore, QBEmax achieved exceptional product purity, with up to 99.4% ± 0.4% of edits consisting of the desired C-to-T conversion, compared to 95.5% ± 2.8% for BE4max [48].
Prime editing systems have also seen substantial improvements through multiple generations. The evolution from PE1 to PE6 has progressively enhanced editing efficiency while minimizing byproducts. In optimized systems using PEmax (an enhanced prime editor) in MMR-deficient cells, researchers achieved remarkably high precise editing rates—reaching ~95% for specific edits with minimal errors by day 28 using engineered pegRNAs (epegRNAs) [19]. A landmark study published in Nature Methods in 2025 established a high-efficiency prime editing platform capable of precise substitution editing with minimal unwanted byproducts. This system demonstrated that in MMR-deficient cell lines, prime editing could achieve 68.9% precise editing of HEK3 +1 T>A and 81.1% of DNMT1 +6 G>C after just 7 days using epegRNAs, with precise editing continuing to increase over time [19].
The critical advantage of prime editing emerges clearly in the comparison of byproduct profiles. Where even advanced base editors like QBEmax still produce low but measurable levels of indels (1.2% ± 0.2%) and impure editing products (0.6%), prime editing systems can achieve near-perfect editing with error rates (including indels and incorrect edits) below 5% for many targets in optimized conditions [19] [48]. This makes prime editing particularly valuable for therapeutic applications where maximum precision is required.
Context-Dependent Performance Variations
The performance of both technologies varies significantly depending on genomic context, cell type, and specific edit being attempted. For base editing, efficiency and byproduct formation are heavily influenced by the sequence context surrounding the target base—specifically the presence of multiple identical bases within the editing window that may lead to bystander edits [8]. Prime editing efficiency shows substantial variation across different genomic loci, with chromatin accessibility and local sequence features affecting success rates [19]. MMR activity presents a particular challenge for prime editing, as cellular repair systems can recognize and reverse prime edits, lowering overall efficiency [19] [13]. To address this, researchers have developed PE4 and PE5 systems that incorporate dominant-negative MLH1 (MLH1dn) to inhibit MMR, significantly boosting editing efficiency from ~20-40% in PE2 to ~50-70% in PE4, and further to ~60-80% in PE5 systems [12].
Table 2: Quantitative Comparison of Byproduct Profiles in Advanced Editing Systems
| Performance Metric | Advanced Base Editing (QBEmax) | Advanced Prime Editing (PEmaxKO + epegRNA) | Notes |
|---|---|---|---|
| Typical Indel Frequency | 1.2% ± 0.2% (average across 17 sites) [48] | <4% (median errors on day 28) [19] | Prime editing indels primarily from imperfect flap resolution |
| Product Purity | 99.4% ± 0.4% desired C-to-T conversion [48] | >95% precise editing for optimized targets [19] | Prime editing purity depends on MMR inhibition |
| Bystander Editing | Significant concern within editing window [3] [8] | Minimal to none [8] [12] | Major differentiator between technologies |
| Off-Target Effects | DNA and RNA off-targets possible from deaminase activity [8] | Minimal off-targets reported [8] [19] | Prime editing shows cleaner off-target profile |
| Theoretical Correction Scope | ~30% of known pathogenic SNVs [3] | ~89% of known pathogenic human genetic variants [3] | Prime editing offers broader therapeutic potential |
Experimental Approaches for Byproduct Assessment
Methodologies for Byproduct Characterization
Rigorous assessment of byproducts requires specialized experimental designs and analytical methods. For comprehensive evaluation of base editing outcomes, the field has adopted deep sequencing approaches that provide quantitative measurements of all editing products at targeted loci. The standard protocol involves amplifying target regions from genomic DNA using PCR, preparing sequencing libraries, and performing high-depth sequencing (typically >10,000x coverage) to detect even low-frequency byproducts [48]. Bioinformatic analysis pipelines then categorize sequencing reads into precise edits, impure edits (including bystander edits and transversions), indels, and unedited sequences. This approach enables precise quantification of product purity—calculated as the proportion of desired base conversions among all editing products [48].
For prime editing, the analytical framework is similar but must account for a wider variety of potential outcomes. The recommended methodology involves targeted amplicon sequencing with unique molecular identifiers (UMIs) to eliminate PCR amplification biases and enable accurate quantification of editing efficiencies and byproducts [19]. The analysis typically categorizes outcomes into three main classes: (1) precise edits containing only the intended modification, (2) outcomes with at least one error (including incorrect nucleotide incorporation or small indels), and (3) unedited sequences [19]. This classification provides a comprehensive view of prime editing fidelity and identifies specific failure modes that need optimization.
Advanced screening approaches have been developed for multiplexed assessment of editing outcomes across many target sites simultaneously. Self-targeting "sensor" libraries that link epegRNA expression cassettes to sensor target sequences enable high-throughput evaluation of prime editing efficiency and specificity across thousands of guide RNA-target pairs [19]. These systems reasonably model editing at endogenous targets while allowing scalable assessment of byproduct formation. For therapeutic applications, additional assays are recommended to assess off-target effects, including whole-genome sequencing or GUIDE-seq for DNA off-targets, and transcriptome-wide sequencing for RNA off-targets particularly relevant for base editors with deaminase domains [8].
Protocol for Systematic Byproduct Evaluation
A robust protocol for comparative assessment of base editing and prime editing byproducts should include the following key steps:
Target Selection: Choose a panel of endogenous genomic sites representing different sequence contexts, chromatin environments, and edit types. Include both optimally and suboptimally positioned targets for each technology.
Editor Delivery: For base editing, deliver editor plasmids or ribonucleoproteins (RNPs) alongside sgRNAs. For prime editing, deliver the prime editor protein or encoding plasmid with specifically designed pegRNAs. Maintain consistent delivery methods and molar ratios for fair comparison.
Harvesting and Analysis Timepoints: Collect samples at multiple timepoints (e.g., 3, 7, 14 days post-editing) to track the evolution of editing outcomes and byproduct accumulation over time.
Sequencing Library Preparation: Extract genomic DNA and amplify target loci with barcoded primers. Use UMI-based approaches to enable accurate quantification and avoid PCR duplicates.
Bioinformatic Analysis: Process sequencing data through established pipelines (e.g., CRISPResso2 for base editing, PrimeEditor for prime editing) to quantify precise edits, byproducts, and indels.
Statistical Comparison: Normalize efficiency measurements and compare byproduct frequencies using appropriate statistical tests, accounting for multiple comparisons where necessary.
This systematic approach enables direct, quantitative comparison of the indel problem and other byproducts across both technologies, providing researchers with actionable data for selecting the optimal editing approach for specific applications.
Visualization of Editing Mechanisms and Byproduct Pathways
The following diagrams illustrate the fundamental mechanisms of base editing and prime editing, highlighting the key steps where byproducts originate.
Diagram 1: Comparative mechanisms of base editing (top) and prime editing (bottom), highlighting key steps where byproducts are generated. Base editing primarily produces bystander edits within its editing window, while prime editing generates minimal byproducts primarily during flap resolution.
Research Reagent Solutions for Byproduct Analysis
Table 3: Essential Research Reagents for Byproduct Characterization Studies
| Reagent Category | Specific Examples | Function in Byproduct Analysis | Considerations for Selection |
|---|---|---|---|
| Editor Systems | PE2, PEmax, PE5, BE4max, QBEmax [12] [48] | Core editing machinery for comparative studies | PE5 includes MLH1dn for MMR inhibition; QBEmax offers higher purity |
| Guide RNA Formats | sgRNA, epegRNA, pegRNA with optimized motifs [8] [19] | Target specification and edit encoding | epegRNAs with tevopreQ1 motif enhance prime editing efficiency [19] |
| Delivery Tools | AAV vectors, lipid nanoparticles, electroporation systems [3] [13] | Intracellular delivery of editing components | Prime editor size complicates AAV packaging; dual AAV systems often needed |
| Analysis Kits | UMI-amplicon sequencing kits, CRISPResso2, PrimeEditor analysis tools [19] | Byproduct quantification and characterization | UMI-based approaches reduce PCR amplification biases |
| Control Systems | Self-targeting sensor libraries, positive/negative control epegRNAs [19] | Experimental normalization and benchmark comparisons | Enable multiplexed assessment across many targets |
| Cell Lines | MMR-deficient variants (e.g., PEmaxKO), HEK293T, K562 [19] [48] | Provide optimized cellular environments | MMR deficiency dramatically improves prime editing efficiency [19] |
The comprehensive comparison of byproduct profiles between base editing and prime editing reveals a clear tradeoff between efficiency and precision. Base editing offers higher efficiency for specific base transitions but carries the burden of bystander edits and low-frequency indels. Prime editing provides exceptional precision with minimal byproducts but faces challenges in efficiency and delivery. The choice between these technologies ultimately depends on the specific application: base editing may be preferable for research applications requiring high efficiency where bystander edits can be tolerated or designed around, while prime editing holds greater promise for therapeutic applications where precision is paramount.
Future directions for both technologies will focus on further minimizing their respective byproduct profiles while maintaining or improving editing efficiency. For base editing, this includes developing editors with narrower activity windows to reduce bystander edits and engineering deaminases with reduced off-target activity [48]. For prime editing, ongoing work focuses on optimizing the pegRNA design, improving reverse transcriptase processivity and fidelity, and developing more efficient delivery strategies for the large editing machinery [8] [19]. As both technologies continue to evolve, their convergence toward a truly optimal editor—one that combines the efficiency of base editing with the precision and versatility of prime editing—represents the next frontier in precision genome editing.
For researchers embarking on genome editing studies, the methodological framework presented here provides a foundation for rigorous byproduct assessment. By implementing standardized protocols for byproduct characterization and utilizing the recommended reagent solutions, the scientific community can generate comparable data across studies and platforms, accelerating the development of ever-more precise genome editing technologies for research and therapeutic applications.
Benchmarking Editing Efficiency and Purity Across Loci
The advent of precision genome editing has revolutionized genetic research and therapeutic development. Among these technologies, prime editing and base editing have emerged as leading tools for making targeted changes without introducing double-strand DNA breaks. This guide provides a systematic comparison of their editing efficiency and purity across various genomic loci, offering researchers and drug development professionals critical data for selecting appropriate editing platforms for specific applications.
While base editors enable efficient single-nucleotide conversions, their editing scope is limited to specific transition mutations and they often produce unwanted bystander edits within their activity window [8] [9]. Prime editing, a more versatile "search-and-replace" technology, theoretically supports all 12 possible base substitutions, small insertions, and deletions with higher precision and fewer off-target effects [25] [12]. However, early prime editing systems suffered from low and variable editing efficiencies that complicated their application in high-throughput functional genomics [19] [49].
Key Performance Metrics: Prime Editing vs. Base Editing
Quantitative Comparison of Editing Performance
Table 1: Benchmarking editing efficiency and purity between optimized prime editing and base editing systems
| Performance Metric | Base Editing | Early Prime Editing (PE2) | Optimized Prime Editing (PEmaxKO + epegRNA) |
|---|---|---|---|
| Editing Scope | 4 transition mutations (C>T, G>A, A>G, T>C) [9] | All 12 point mutations, small insertions, deletions [25] | All 12 point mutations, small insertions, deletions [19] |
| Typical Efficiency Range | 50-80% for compatible edits [9] | 1-20% (highly variable) [49] | 20-95% (median ~80% in optimized systems) [19] |
| Purity (Intended Edit %) | Moderate (bystander editing common) [8] [9] | High (minimal byproducts) [25] | Very high (94.9% precise editing demonstrated) [19] |
| DSB Formation | No [9] | No [25] | No [19] |
| Bystander Editing | Yes (within 4-5nt window) [8] [9] | Minimal [25] | Minimal [19] |
| MMR Interference | Not applicable | High inhibition of small edits [19] | Minimal (in MMR-deficient systems) [19] |
| Key Limitations | Restricted editing scope, bystander mutations, off-target deamination [8] [9] | Low efficiency, MMR sensitivity [19] [25] | Large construct size, delivery challenges [13] |
Locus-to-Locus Variability in Editing Efficiency
Editing efficiency varies significantly across genomic loci for both base editing and prime editing technologies. In a benchmark study using a self-targeting library of 2,000 epegRNA-target pairs, prime editing efficiency in MMR-deficient (PEmaxKO) cells showed remarkable variation [19]. By day 28 post-transduction, 75.5% (1,453/1,974) of edits reached ≥75% precise editing in PEmaxKO cells, compared to only 20.2% (388/1,974) in MMR-proficient (PEmax) cells [19]. This demonstrates that while optimized prime editing systems can achieve high efficiency at many loci, MMR status dramatically influences outcomes.
For base editing, efficiency also varies across targets but generally remains higher than early prime editing systems. However, this efficiency comes at the cost of precision. Base editors frequently modify non-target bases within their activity window, with bystander editing observed in up to 70% of editing events depending on the sequence context [9]. This significantly reduces the purity of intended edits compared to prime editing systems.
Table 2: Locus-specific editing efficiency comparison between base editing and prime editing
| Genomic Locus | Edit Type | Base Editing Efficiency | Prime Editing Efficiency | Notes |
|---|---|---|---|---|
| HEK3 +1 | T>A | Not possible with BE | 94.9% (PEmaxKO) vs 7.8% (PEmax) [19] | Demonstrates MMR sensitivity |
| DNMT1 +6 | G>C | Not possible with BE | 91.8% (PEmaxKO) vs 78.7% (PEmax) [19] | G>C edits evade MMR to some degree |
| OsCDC48 | Not specified | ~60-80% (typical for compatible edits) | 29.17% [17] | Plant system example |
| OsACC1 | Not specified | ~50-70% (typical for compatible edits) | 0% [17] | Plant system with no detected editing |
| Various (2,000 targets) | +5 G>H (A,T,C) | 50-80% for compatible edits | 75.5% reached ≥75% efficiency (PEmaxKO) [19] | High-throughput sensor library |
Experimental Protocols for Benchmarking
High-Efficiency Prime Editing Workflow
The following workflow was established to benchmark prime editing efficiency and purity across multiple loci [19] [49]:
Cell Line Engineering
- Generate clonal cell lines (e.g., K562) constitutively expressing optimized prime editor (PEmax) from AAVS1 safe-harbor locus with co-expressed EGFP reporter
- Create MMR-deficient derivative (PEmaxKO) by knocking out MLH1 gene
- Validate editor expression and MMR status via Western blot and functional assays
epegRNA Library Design and Delivery
- Design epegRNAs with 3' tevopreQ1 motif to enhance stability
- Include 10-15nt primer binding site (PBS) and ~25-40nt reverse transcription template (RTT)
- Clone library into lentiviral vectors with ~240,000 epegRNAs targeting ~17,000 codons
- Transduce at low MOI (0.7) to ensure single integration events
- Select transduced cells with appropriate antibiotics
Editing Quantification and Analysis
- Harvest cells at multiple timepoints (7, 14, 21, 28 days post-transduction)
- Amplify and sequence target loci using high-throughput sequencing
- Analyze outcomes with custom pipeline categorizing: (1) precise edits only, (2) edits with ≥1 error, (3) unedited sequence
- Calculate enrichment/depletion of epegRNAs to determine functional impacts
Base Editing Benchmarking Protocol
Editor Delivery and Expression
- Deliver base editor (CBE or ABE) via lentiviral transduction or transient transfection
- Include guide RNA targeting specific loci with compatible PAM sequences
- Validate editor expression and nuclear localization
Editing Assessment
- Harvest cells 72-96 hours post-delivery
- Extract genomic DNA and amplify target regions
- Sequence using next-generation sequencing
- Quantify efficiency as percentage of reads with intended base conversion
- Calculate bystander editing rate as percentage of edits with additional unintended changes within activity window
Molecular Mechanisms Underlying Performance Differences
Prime Editing Mechanism
Prime editing operates through a complex multi-step mechanism that enables precise "search-and-replace" genome editing without double-strand breaks [8] [25]:
- Target Recognition: The prime editor (nCas9-H840A fused to reverse transcriptase) binds to target DNA directed by the pegRNA spacer sequence
- DNA Nicking: nCas9 creates a single-strand nick in the DNA complementary to the pegRNA
- Reverse Transcription: The nicked DNA primes reverse transcription using the pegRNA's RT template, synthesizing new DNA containing the desired edit
- Flap Resolution: Cellular enzymes resolve the resulting DNA flap structure, incorporating the edited strand
- Complementary Strand Editing (PE3): An additional sgRNA directs nicking of the non-edited strand to promote permanent integration of the edit
Base Editing Mechanism
Base editing employs a simpler but less precise mechanism [8] [9]:
- Target Binding: Cas9 nickase-deaminase fusion binds DNA directed by sgRNA
- R-Loop Formation: Cas9 displaces non-target strand, creating single-stranded DNA bubble
- Deamination: Deaminase enzyme (APOBEC for CBEs, evolved TadA for ABEs) chemically converts bases within the activity window (typically 4-5 nucleotides)
- DNA Repair: Cellular mismatch repair processes complete the base conversion, with UGI domain (in CBEs) preventing base excision repair
- Outcome: Permanent point mutation without double-strand break, but with potential bystander edits
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key research reagents for benchmarking editing technologies
| Reagent Category | Specific Examples | Function in Experiment | Considerations |
|---|---|---|---|
| Editor Systems | PEmax, PE2, PE3, PE5 [19] [12] | Catalyze the desired DNA edits | PEmax shows improved efficiency over PE2; PE5 incorporates MLH1dn to inhibit MMR |
| Guide RNA Formats | epegRNA (with tevopreQ1), standard pegRNA [19] [13] | Specify target site and template edit | epegRNAs show 3-4x higher efficiency due to improved stability |
| Cell Lines | PEmax-K562, PEmaxKO (MLH1-/-) [19] | Provide cellular context for editing | MMR-deficient lines dramatically improve prime editing efficiency for small edits |
| Delivery Vectors | Lentiviral vectors, lipid nanoparticles [13] | Introduce editing components into cells | Lentiviral enables stable integration; size constraints for packaging large PEs |
| MMR Inhibitors | MLH1dn (dominant-negative) [12] | Enhance prime editing efficiency | Particularly important for small edits highly susceptible to MMR |
| Selection Markers | Puromycin, GFP [19] | Enrich for successfully transduced cells | Critical for pooled screens to maintain library representation |
| Sequencing Tools | NGS platforms, custom analysis pipelines [19] | Quantify editing outcomes and efficiency | Essential for characterizing efficiency and purity across loci |
The benchmarking data presented demonstrates that while both base editing and prime editing enable precise genome editing without double-strand breaks, they exhibit distinct performance characteristics across different genomic loci. Base editing offers higher efficiencies for compatible edits but suffers from limited scope and unwanted bystander mutations. Prime editing, particularly when optimized with MMR deficiency and epegRNAs, achieves remarkable precision and broader editing scope, with recent systems reaching >90% efficiency at many loci.
The choice between these technologies depends on the specific research requirements: base editing may be preferable for straightforward transition mutations where high efficiency is paramount, while prime editing excels when precision, versatility, or transversion mutations are required. As both technologies continue to evolve, researchers now have multiple powerful options for precision genome editing across diverse genomic contexts.
In the realm of therapeutic genome editing, the transition from conventional CRISPR-Cas nucleases to more advanced precision editors represents a paradigm shift toward greater control over genetic outcomes. While traditional nuclease-based approaches rely on double-strand breaks (DSBs) and error-prone repair processes—often resulting in a complex mixture of insertion and deletion (indel) byproducts—newer technologies aim to minimize these unpredictable consequences [12] [9]. Among the most critical metrics for evaluating these advanced editors is the edit:indel ratio, which quantifies the frequency of desired precise edits relative to unwanted indels. This ratio serves as a crucial indicator of editing purity, with profound implications for therapeutic safety [40] [50].
The emergence of base editing and prime editing technologies has fundamentally changed the landscape of precision genome engineering. Base editors, employing deaminase enzymes fused to Cas9 nickase, enable single nucleotide conversions without creating DSBs, but they operate within constrained editing windows and can produce bystander edits at neighboring bases [12] [9]. Prime editors, utilizing a Cas9 nickase-reverse transcriptase fusion guided by a specialized pegRNA, offer a more versatile "search-and-replace" capability for installing a broader range of genetic changes while similarly avoiding DSBs [12] [39]. For both platforms, the edit:indel ratio provides an essential quantitative measure of their precision, directly impacting their therapeutic viability and safety profile, particularly as these technologies advance toward clinical applications [40] [50].
Technology Comparison: Base Editing vs. Prime Editing
Fundamental Mechanisms and Editing Capabilities
Base editors and prime editors represent distinct technological approaches to precision genome editing, each with characteristic mechanisms, capabilities, and limitations. Understanding their fundamental differences is essential for appreciating how edit:indel ratios are contextualized within each system.
Base editing employs a catalytically impaired Cas9 (nickase) fused to a nucleobase deaminase enzyme, which directly converts one DNA base to another without breaking both DNA strands [9]. Cytosine base editors (CBEs) facilitate C-to-T conversions, while adenine base editors (ABEs) enable A-to-G changes [41]. The editing occurs within a defined "editing window" of approximately 4-5 nucleotides, which can lead to bystander edits when multiple targetable bases are present within this window [12] [9]. While base editors typically produce very few indels due to their avoidance of double-strand breaks, their primary precision challenge lies in controlling which bases within the editing window are modified, rather than preventing indel formation [9].
Prime editing utilizes a more complex mechanism involving a Cas9 nickase-reverse transcriptase fusion protein programmed with a prime editing guide RNA (pegRNA) [12] [39]. The pegRNA both directs the editor to the target site and encodes the desired edit. After nicking the target DNA strand, the reverse transcriptase uses the pegRNA template to synthesize DNA containing the desired sequence change, which is then incorporated into the genome [39]. This "search-and-replace" mechanism enables a wider variety of edits—including all 12 possible base-to-base conversions, small insertions, and deletions—without being confined to a narrow editing window [12] [39]. However, the multi-step process of prime editing introduces more opportunities for competing cellular repair pathways to generate indel byproducts, making edit:indel ratios a particularly critical metric for this technology [40].
Direct Performance Comparison
The table below summarizes the key characteristics of both editing approaches, highlighting factors that influence their precision and therapeutic potential:
Table 1: Comparison of Base Editing and Prime Editing Technologies
| Feature | Base Editing | Prime Editing |
|---|---|---|
| Core Mechanism | Deaminase-mediated base conversion | Reverse transcriptase-mediated DNA synthesis |
| DNA Break Type | Single-strand nick | Single-strand nick |
| Editing Scope | C-to-T or A-to-G conversions | All 12 base conversions, insertions, deletions |
| Editing Window | Narrow (4-5 nucleotides) | Broad (extended template) |
| Primary Precision Challenge | Bystander edits within window | Competing flap resolution leading to indels |
| Template Requirement | None | pegRNA-encoded template |
| Therapeutic Applications | Single nucleotide polymorphisms | Diverse mutations including insertions, deletions |
Quantifying Precision: Edit:Indel Ratio Fundamentals
Experimental Measurement and Significance
The edit:indel ratio is determined through targeted deep sequencing of edited genomic regions, comparing the frequency of precisely edited alleles to those containing insertions or deletions [40]. This metric has emerged as a paramount indicator of editing purity, with profound implications for therapeutic development. Unlike editing efficiency alone, which measures how frequently desired edits occur, the edit:indel ratio quantifies the purity of the editing outcome—the proportion of edited cells that contain the precise intended modification rather than potentially deleterious indels [40] [50].
In therapeutic contexts, high indel rates pose significant safety concerns, as these unpredictable mutations can disrupt essential genes, activate oncogenes, or inactivate tumor suppressors [40] [50]. Consequently, achieving high edit:indel ratios is particularly crucial for clinical applications. Recent advances in prime editing have demonstrated remarkable improvements in this metric, with next-generation editors achieving ratios as high as 543:1—meaning for every 543 desired edits, only one indel occurs [40] [50]. This represents a dramatic improvement over earlier systems, where ratios could be as low as 7:1, indicating one indel for every seven desired edits [50].
Progression of Prime Editor Evolution
The development of prime editing systems has followed an iterative path of optimization, with successive generations demonstrating improved editing efficiency and reduced indel formation. The following diagram illustrates the key stages in the evolution of prime editors and their associated improvements:
Figure 1: The evolution of prime editors from initial proof-of-concept (PE1) to highly precise systems (vPE) through multiple optimization strategies.
Table 2: Evolution of Prime Editing Systems and Their Characteristics
| Editor | Key Innovation | Typical Editing Efficiency | Edit:Indel Ratio | Reference |
|---|---|---|---|---|
| PE1 | Foundational system | ~10-20% | Not characterized | [12] |
| PE2 | Optimized reverse transcriptase | ~20-40% | Not characterized | [12] |
| PE3/PE3b | Additional nicking sgRNA | ~30-50% | Low (indels increased) | [12] [39] |
| PE4/PE5 | MMR inhibition (MLH1dn) | ~50-70% | Improved | [12] [39] |
| PEmax | Optimized architecture | Similar to PE2-PE5 | Variable | [39] |
| PE6a-g | Evolved RT variants | ~70-90% | Improved for specific edits | [12] [39] |
| PE7 | La protein fusion for pegRNA stability | ~80-95% | Enhanced | [12] [39] |
| vPE | Nick-relaxing Cas9 mutations | High efficiency maintained | Up to 543:1 | [40] [50] |
Breakthrough: Next-Generation Prime Editors with Minimal Errors
The vPE System: Mechanism and Performance
The most recent breakthrough in minimizing indel formation comes from the development of the very-precise Prime Editor (vPE), which incorporates strategic mutations in the Cas9 nickase to relax nick positioning and promote degradation of the competing 5' DNA strand [40] [51] [50]. This approach addresses a fundamental limitation in previous prime editing systems: the bias toward retaining the original unedited DNA strand during flap resolution, which could lead to alternative repair pathways generating indels [40].
The vPE system builds upon earlier innovations including the precise Prime Editor (pPE) and extra-precise Prime Editor (xPE), incorporating error-suppressing Cas9 mutations into the PE7 framework [51]. By engineering Cas9 variants with mutations such as K848A and H982A, researchers destabilized the binding of the non-target strand 5' end, making it more susceptible to cellular nucleases [40]. This degradation of the competing original strand favors the incorporation of the newly synthesized edited strand, significantly reducing opportunities for indel formation during the repair process [40] [50].
The performance metrics of vPE represent a substantial advancement in editing precision. Compared to previous systems like PEmax, vPE reduces indel errors by up to 60-fold while maintaining high editing efficiency, resulting in edit:indel ratios as high as 543:1 [40] [50]. This means that for every 543 desired edits, only a single indel occurs—dramatically higher than the approximately 7:1 ratio observed with earlier systems in their most-used editing mode [50]. This level of precision marks a critical milestone for therapeutic applications where minimizing unintended genetic alterations is paramount for safety.
Comparative Performance Data
The table below summarizes quantitative comparisons between editing systems, demonstrating the progressive improvement in edit:indel ratios:
Table 3: Comparative Performance of Advanced Prime Editing Systems
| Editing System | Editing Mode | Average Editing Efficiency | Indel Frequency | Edit:Indel Ratio |
|---|---|---|---|---|
| PEmax | pegRNA + ngRNA | Baseline | 1 in 7 edits | ~7:1 |
| PEmax | High-precision | Lower efficiency | 1 in 122 edits | ~122:1 |
| pPE | pegRNA + ngRNA | Maintained | 7.6-fold reduction vs PEmax | ~47:1 |
| xPE | pegRNA + ngRNA | Maintained | Further reduction | Improved |
| vPE | pegRNA + ngRNA | High efficiency maintained | 1 in 543 edits | ~543:1 |
| vPE | Various loci tested | Comparable to PEmax | 26-fold reduction vs PEmax | Up to 543:1 |
Experimental Protocols for Assessing Edit:Indel Ratios
Methodology for Evaluating Editing Precision
Rigorous assessment of edit:indel ratios requires standardized experimental approaches that reliably quantify both desired edits and unwanted byproducts. The following protocol outlines key methodological considerations based on recent landmark studies:
5.1.1 Cell Culture and Editor Delivery
- Cell Lines: Experiments are typically conducted in human embryonic kidney (HEK293T) cells, with validation in other therapeutically relevant cell types [40].
- Delivery Methods: Prime editor components can be delivered via plasmid transfection, viral vectors (e.g., lentivirus, AAV), or as ribonucleoprotein (RNP) complexes [40] [39].
- Dosage Optimization: Titration of editor-to-pegRNA ratios is critical for maximizing edit:indel ratios while maintaining efficiency [40].
5.1.2 Target Selection and pegRNA Design
- Locus Selection: Evaluation across multiple genomic loci (e.g., CXCR4, EMX1, TGFB1, KRAS) with varying sequence contexts and chromatin states [40].
- pegRNA Design: Utilizing engineered pegRNAs (epegRNAs) with structured RNA motifs (e.g., evopreQ, mpknot) at the 3' end to protect against degradation and improve editing efficiency [12] [39].
- Nicking gRNA Design: For PE3/PE5 systems, designing nicking gRNAs with appropriate spacing from the pegRNA nick site to minimize double-strand break formation [12] [39].
5.1.3 Analysis and Quantification
- Amplicon Sequencing: Targeted deep sequencing (150-200bp amplicons) of edited genomic regions with minimum 10,000x read depth [40].
- Bioinformatic Analysis: Computational pipelines to distinguish precise edits from indels and other byproducts, with careful filtering of sequencing artifacts [40].
- Edit:Indel Calculation: Ratio determination by dividing the frequency of precise edits by the frequency of indels at the target site [40] [50].
The following workflow diagram illustrates the key experimental steps in evaluating edit:indel ratios:
Figure 2: Key experimental steps for determining edit:indel ratios, from editor design to final quantification.
Essential Research Reagents and Tools
The table below outlines critical experimental components required for conducting precision editing evaluations:
Table 4: Essential Research Reagents for Precision Editing Studies
| Reagent Category | Specific Examples | Function/Purpose | Key Considerations |
|---|---|---|---|
| Editor Plasmids | PEmax, PE6, PE7, vPE variants | Encodes the prime editor protein | Codon optimization, nuclear localization signals |
| pegRNA Expression Systems | epegRNA vectors with RNA stability motifs | Guides editing and templates new sequence | 3' pseudoknot structures for degradation protection |
| Delivery Tools | Lentiviral vectors, AAV, lipid nanoparticles | Cellular delivery of editing components | Size constraints, especially for AAV packaging |
| MMR Inhibitors | MLH1dn (dominant-negative) | Temporary mismatch repair inhibition | Improves editing efficiency in PE4/PE5 systems |
| Analysis Reagents | PCR primers for target amplification, sequencing libraries | Amplification and sequencing of edited loci | Minimum 10,000x read depth recommended |
| Control Elements | Non-targeting pegRNAs, untreated cells | Experimental controls for background mutation rates | Essential for distinguishing editor-specific effects |
Implications for Therapeutic Development
The remarkable improvement in edit:indel ratios achieved by next-generation prime editors has profound implications for therapeutic genome editing. The transition from early prime editors with concerning indel rates to systems like vPE with ratios exceeding 500:1 significantly enhances the safety profile of these technologies [40] [50]. This level of precision reduces the risk of introducing potentially oncogenic mutations through editing byproducts, addressing a key regulatory concern for gene therapies.
Clinical applications of precision editing are already advancing rapidly. The first base-edited cell therapy achieved remission in a 13-year-old girl with relapsed T-cell leukemia, demonstrating the therapeutic potential of these technologies [41]. For prime editing, the first clinical trial was approved in 2024 for chronic granulomatous disease, a genetic immune deficiency [41]. As these therapies progress, edit:indel ratios will serve as a critical quality control metric and regulatory benchmark, ensuring that therapeutic editing meets the stringent safety standards required for clinical use [40] [50].
Future developments will likely focus on further optimizing edit:indel ratios while maintaining high efficiency across diverse genomic contexts and cell types. Additionally, addressing challenges such as delivery efficiency, immunogenicity, and tissue-specific optimization will be essential for realizing the full therapeutic potential of these precision editing platforms [13] [39]. The continued refinement of edit:indel ratios represents a crucial step toward safe, effective, and broadly applicable genetic medicines.
The advent of precision genome editing has revolutionized biomedical research and therapeutic development, moving beyond the disruptive capabilities of early CRISPR-Cas nuclease systems. Among the most significant advancements are base editing and prime editing, two technologies that enable precise genetic modifications without requiring double-stranded DNA breaks (DSBs) [9]. While both represent substantial progress over conventional CRISPR-Cas9 systems, they differ fundamentally in their mechanisms, capabilities, and limitations. Base editing, developed first, utilizes deaminase enzymes to directly convert one base to another but remains restricted to specific transition mutations [13]. Prime editing, a more recent innovation, employs a reverse transcriptase to write new genetic information directly into the genome, offering substantially broader editing capabilities [12] [8]. This comparative analysis examines the editing scope, flexibility, and limitations of these two precision editing platforms, providing researchers with a framework for selecting the appropriate technology for specific experimental or therapeutic applications.
Molecular Mechanisms and Editing Scope
Base Editing Architecture and Capabilities
Base editors are fusion proteins consisting of a catalytically impaired Cas protein (nickase) tethered to a nucleotide deaminase enzyme [9]. Two primary classes have been developed: Cytosine Base Editors (CBEs) convert cytosine to thymine (C→T), while Adenine Base Editors (ABEs) convert adenine to guanine (A→G) [13] [9]. The editing process occurs within a well-defined "editing window" of approximately 4-5 nucleotides in the spacer region, with peak activity typically observed at positions 4-8 in the protospacer [26]. This restricted activity window represents a significant limitation, as it often leads to unwanted "bystander edits" where adjacent nucleotides within the window are unintentionally modified [26]. Base editors are further constrained by protospacer adjacent motif (PAM) requirements and cannot perform transversion mutations, insertions, or deletions [12] [9].
Table 1: Base Editing Technical Specifications
| Characteristic | Cytosine Base Editors (CBEs) | Adenine Base Editors (ABEs) |
|---|---|---|
| Core Mechanism | Cytidine deaminase + UGI domain | Engineered tRNA adenosine deaminase |
| Base Conversion | C•G to T•A | A•T to G•C |
| Editing Window | 4-5 nucleotides | 4-5 nucleotides |
| Primary Components | nickase Cas9-APOBEC1-UGI | nickase Cas9-TadA |
| Efficiency Range | Up to 92% specificity (BE4-Gam) [26] | Up to 97% specificity (ABE7.10) [26] |
| Key Limitations | Bystander edits, C→T restrictions | Bystander edits, A→G restrictions |
Prime Editing Architecture and Capabilities
Prime editing represents a more versatile approach that overcomes many of base editing's limitations. The system comprises a Cas9 nickase (H840A) fused to an engineered reverse transcriptase (RT) from Moloney Murine Leukemia Virus (M-MLV), programmed with a specialized prime editing guide RNA (pegRNA) [12] [8] [13]. The pegRNA not only directs the complex to the target site but also encodes the desired edit within its reverse transcription template (RTT) region [13]. This innovative architecture enables prime editing to perform all 12 possible base-to-base conversions, in addition to targeted insertions and deletions without requiring donor DNA templates or creating double-strand breaks [12] [8] [13]. The editing process occurs through a multi-step mechanism: target recognition and nicking, primer binding, reverse transcription, flap equilibration, and repair-mediated incorporation of the edit [13] [32].
Prime Editing Mechanism: A visual representation of the multi-step prime editing process, from target recognition to edit incorporation.
The evolution of prime editors from PE1 to PE7 has progressively addressed initial limitations in efficiency and specificity. PE2 introduced optimized reverse transcriptase, while PE3 added a second nicking guide RNA to enhance editing efficiency by encouraging cellular repair machinery to use the edited strand as a template [12] [8]. Later versions (PE4-PE7) incorporated additional refinements such as mismatch repair inhibition (MLH1dn) and pegRNA stabilization strategies to further improve performance [12].
Table 2: Prime Editing System Evolution
| Editor Version | Key Components | Editing Efficiency | Major Innovations |
|---|---|---|---|
| PE1 | nCas9(H840A)-RT, pegRNA | ~10-20% | Proof-of-concept system |
| PE2 | Optimized RT, pegRNA | ~20-40% | Enhanced RT processivity and stability |
| PE3 | RT, pegRNA, additional sgRNA | ~30-50% | Dual nicking strategy for enhanced efficiency |
| PE4 | RT, pegRNA, MLH1dn | ~50-70% | Mismatch repair inhibition |
| PE5 | RT, pegRNA, additional sgRNA, MLH1dn | ~60-80% | Combined MMR inhibition and strand nicking |
| PE6 | Compact RT variants, epegRNAs | ~70-90% | Improved delivery and pegRNA stability |
| PE7 | Modified RT, epegRNAs, La protein | ~80-95% | Enhanced pegRNA stability in challenging cells |
Precision and Safety Profiles
Off-Target Effects and Editing Purity
The precision of genome editing tools is paramount for therapeutic applications. Base editors, while generally producing fewer indels than nuclease-based approaches, suffer from significant bystander editing problems where multiple nucleotides within the editing window are modified [26]. Recent deep learning models (CRISPRon-ABE and CRISPRon-CBE) have been developed to better predict these outcomes, revealing that base editors maintain "a large editing window, which can introduce unintended bystander edits" [26]. Additionally, the deaminase enzymes in base editors (APOBEC and TadA) can exhibit off-target activity on both DNA and RNA, creating potential safety concerns for clinical applications [12] [8].
Prime editing demonstrates superior editing purity with significantly reduced bystander effects and minimal off-target activity [12] [8]. The requirement for three independent hybridization events (spacer binding, PBS annealing, and flap resolution) enhances specificity, though early versions utilizing the H840A nickase could inadvertently generate double-strand breaks [8]. This limitation has been addressed in newer systems through additional mutations (N863A) that further reduce off-target and on-target DSBs [8]. A comparative study assessing editing outcomes across multiple cell lines reported that optimized prime editing systems can achieve up to 80% editing efficiency with high precision [52].
Practical Performance Comparison
Table 3: Direct Comparison of Editing Technologies
| Parameter | Base Editing | Prime Editing |
|---|---|---|
| DSB Formation | Minimal | None |
| Theoretical Editing Scope | 4 transition mutations | All 12 point mutations, insertions, deletions |
| Bystander Edits | Common challenge | Greatly reduced |
| Editing Efficiency Range | 30-97% (varies by editor) [26] | 10-95% (version-dependent) [12] [52] |
| PAM Restrictions | Yes | Yes, but expanded by Cas variants |
| Delivery Challenges | Moderate (size ~5-6kb) | Significant (size ~6.5kb+) |
| Therapeutic Validation | Multiple in vivo models [7] | Emerging preclinical data |
Experimental Optimization and Implementation
Efficiency Enhancement Strategies
Both editing platforms require optimization to achieve maximal performance. Base editing efficiency is influenced by sequence context, Cas9 activity at the target site, and the specific deaminase variant employed [26]. Computational tools have become essential for designing optimal base editing experiments, with multi-dataset training approaches significantly improving prediction accuracy for both editing efficiency and bystander outcomes [26].
Prime editing efficiency is enhanced through several strategies: engineered pegRNAs (epegRNAs) incorporating structured RNA motifs (evopreQ, mpknot) at the 3' end protect against degradation and improve editing efficiency by 3-4 fold [8]. The recently developed proPE system extends the editing window and enhances efficiency where standard prime editing performs poorly by using a second non-cleaving sgRNA to target the reverse transcriptase template near the edit site [32]. Systematically optimized delivery methods, including stable genomic integration via the piggyBac transposon system combined with lentiviral delivery of pegRNAs, have achieved editing efficiencies up to 80% across multiple cell lines [52].
Delivery Considerations
The substantial size of prime editing components presents notable delivery challenges, particularly for viral vectors with limited packaging capacity. The split prime editor (sPE) system addresses this by separating nCas9 and RT into independently functioning units, enabling delivery via dual AAV vectors [8]. This approach has successfully edited the β-catenin gene in mouse liver and corrected mutations in a type I tyrosinemia model [8]. Base editors, while smaller than prime editors, still require sophisticated delivery systems, with most in vivo studies utilizing split-intein dual AAV vectors or emerging lipid nanoparticle (LNP) approaches [7].
Research Reagent Solutions
Table 4: Essential Research Tools for Precision Gene Editing
| Reagent/Tool | Function | Application Notes |
|---|---|---|
| pegRNA | Target specification and edit template | 120-145 nt length; requires stabilization motifs [13] |
| epegRNA | Enhanced stability pegRNA | 3-4 fold efficiency improvement with 3' structure motifs [8] |
| Prime Editor Plasmids | PE2, PEmax, PE5 systems | Available from Addgene (#132775, #174828) [52] |
| PiggyBac Transposon | Stable genomic integration | Enables sustained editor expression [52] |
| CRISPRon-ABE/CBE | Deep learning prediction tools | Web server for gRNA design optimization [26] |
| Dual AAV Vectors | In vivo delivery | Necessary for larger editors; split-intein systems [8] [7] |
| MLH1dn | Mismatch repair inhibition | Enhances editing persistence (PE4/PE5 systems) [12] |
Base editing and prime editing represent complementary technologies in the precision genome editing landscape, each with distinct advantages and limitations. Base editors offer higher efficiency for specific transition mutations and have demonstrated robust therapeutic efficacy in multiple disease models [7]. Prime editors provide substantially greater versatility, enabling a broader range of genetic modifications with superior editing purity, though currently with more variable efficiency and greater delivery challenges [12] [8].
The choice between these technologies depends heavily on the specific research or therapeutic objective. For defined transition mutations (C→T or A→G) where high efficiency is paramount and bystander edits can be managed or are absent, base editing remains the preferable option. For more complex edits including transversions, insertions, deletions, or in contexts where maximal precision is required, prime editing offers unique capabilities. As both technologies continue to evolve through protein engineering, improved delivery strategies, and enhanced computational design tools, their complementary applications will expand, further enabling the precise genetic manipulations necessary for advanced biomedical research and therapeutic development.
Conclusion
The choice between base editing and prime editing is not a matter of one technology being superior, but rather of selecting the right tool for the specific genetic modification goal. Base editors excel in efficiently installing specific transition mutations but are constrained by their limited editing scope and potential for bystander edits. Prime editing, while historically challenged by variable efficiency, offers unparalleled versatility for installing a broad range of precise changes with minimal byproducts, a advantage solidified by next-generation editors like vPE that achieve remarkably high edit-to-indel ratios. Future directions will focus on further improving the efficiency and delivery of these tools, expanding the targetable genomic landscape, and moving promising preclinical results into clinical applications. For researchers and drug developers, this evolving toolkit promises to unlock new frontiers in functional genomics, disease modeling, and the development of transformative genetic therapies.
References
- 1. Base editing: precision chemistry on the genome and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6535181/]
- 2. Back to Basics - Base & Prime Editing [https://frontlinegenomics.com/back-to-basics-base-and-prime-editing/]
- 3. The Arrival of Precision Gene Therapy with Base and ... [https://hp-ne.com/blog/the-arrival-of-precision-gene-therapy-with-base-and-prime-editing/]
- 4. CRISPR 101: Cytosine and Adenine Base Editors [https://blog.addgene.org/single-base-editing-with-crispr]
- 5. Base editing: advances and therapeutic opportunities - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC7721651/]
- 6. Deep learning models simultaneously trained on multiple ... [https://www.nature.com/articles/s41467-025-65200-5]
- 7. In vivo precision base editing to rescue mouse models of ... [https://www.sciencedirect.com/science/article/pii/S2162253125001763]
- 8. Emerging trends in prime editing for precision genome ... [https://www.nature.com/articles/s12276-025-01463-8]
- 9. cutting-edge applications of base editing and prime editing in ... [https://translational-medicine.biomedcentral.com/articles/10.1186/s12967-024-05957-3]
- 10. Improvement of base editors and prime editors advances ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8968349/]
- 11. Base editing and prime editing in laboratory animals - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC8918876/]
- 12. Emerging trends in prime editing for precision genome ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12322275/]
- 13. Prime Editing as a Precision Gene Editing Tool [https://www.synthego.com/guide/crispr-methods/prime-editing/]
- 14. Prime editing: therapeutic advances and mechanistic insights [https://www.nature.com/articles/s41434-024-00499-1]
- 15. MIT Researchers Set New Benchmark for Genome Editing ... [https://joshuaberkowitz.us/blog/news-1/mit-researchers-set-new-benchmark-for-genome-editing-precision-with-prime-editing-breakthrough-1166]
- 16. Prime Editing: The Next Frontier in Precision Gene Therapy [https://pubmed.ncbi.nlm.nih.gov/41038616/]
- 17. Prime Editing for Crop Improvement: A Systematic Review ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12385346/]
- 18. Systematic optimization of prime editing for enhanced ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12069386/]
- 19. A benchmarked, high-efficiency prime editing platform for ... [https://www.nature.com/articles/s41592-024-02502-4]
- 20. High-throughput evaluation of genetic variants with prime ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12520993/]
- 21. Deep learning models simultaneously trained on multiple ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12595020/]
- 22. EXPERT expands prime editing efficiency and range of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11822059/]
- 23. Systematic optimization of prime editing for the efficient ... [https://www.nature.com/articles/s41551-024-01233-3]
- 24. Optimised genome editing for precise DNA insertion and ... [https://elifesciences.org/reviewed-preprints/107475]
- 25. Prime editing for precise and highly versatile genome ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10989687/]
- 26. Breaking: Deep learning multi-dataset training improves ... [https://crisprmedicinenews.com/news/breaking-deep-learning-multi-dataset-training-improves-crispr-base-editing-predictions/]
- 27. Mass screening of genetic variants can clarify disease risk | Crick [https://www.crick.ac.uk/news/2025-03-21_mass-screening-of-genetic-variants-can-clarify-disease-risk]
- 28. variant interpretation using CRISPR Saturation prime editing [https://www.nature.com/articles/s41587-021-01201-1?error=cookies_not_supported&code=74a0cea1-ecbc-4edb-9141-9ebf1013c7d7]
- 29. High throughput PRIME identify editing DNA... screens functional [https://pmc.ncbi.nlm.nih.gov/articles/PMC10370011/]
- 30. CRISPR Agriculture News Today: 7 Powerful Advances in ... [https://farmonaut.com/news/crispr-agriculture-news-today-7-powerful-advances-in-2025]
- 31. Progress and Pressure: Taking Stock of the CRISPR Landscape [https://www.medscape.com/viewarticle/progress-and-pressure-taking-stock-crispr-landscape-2025a1000rzr]
- 32. ProPE expands the prime editing window and enhances ... [https://www.nature.com/articles/s41929-025-01406-6]
- 33. CRISPR-Cas9 DNA Base-Editing and Prime-Editing - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC7503568/]
- 34. Bystander base editing interferes with visual function ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11526940/]
- 35. CRISPR editors gain precision via evolved hotspots [https://crisprmedicinenews.com/news/crispr-editors-gain-precision-via-evolved-hotspots/]
- 36. Spectrum of DNA mismatch repair failures viewed through the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8919091/]
- 37. Prime editing efficiency and fidelity are enhanced in the ... [https://www.nature.com/articles/s41467-022-28442-1]
- 38. Current Status and Challenges of DNA Base Editing Tools [https://pmc.ncbi.nlm.nih.gov/articles/PMC7474268/]
- 39. Prime Editing: Adding Precision and Flexibility to CRISPR ... [https://blog.addgene.org/prime-editing-crisp-cas-reverse-transcriptase]
- 40. Engineered prime editors with minimal genomic errors [https://www.nature.com/articles/s41586-025-09537-3]
- 41. David Liu Wins 2025 Breakthrough Prize for Base Editing and Prime ... [https://www.the-scientist.com/david-liu-wins-2025-breakthrough-prize-for-base-editing-and-prime-editing-72888]
- 42. Optimized Ribonucleoprotein Complexes Enhance Prime ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12345536/]
- 43. A more precise way to edit the genome | MIT News | Massachusetts... [https://news.mit.edu/2025/more-precise-way-edit-genome-0917]
- 44. MIT Scientists Unlock a New Level of Precision in Gene ... [https://scitechdaily.com/mit-scientists-unlock-a-new-level-of-precision-in-gene-editing/]
- 45. Determining Performance Metrics for Targeted Next ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6172655/]
- 46. Comparison of the performance of an amplicon sequencing ... [https://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-020-6557-5]
- 47. Development of a novel amplicon based whole-genome ... [https://www.nature.com/articles/s42003-025-08159-9]
- 48. QBEmax is a sequence-permuted and internally protected ... [https://www.nature.com/articles/s41587-025-02641-9]
- 49. A benchmarked, high-efficiency prime editing platform for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10996517/]
- 50. Modified Prime Genomic Editing Greatly Improves Precision [https://www.insideprecisionmedicine.com/topics/precision-medicine/modified-prime-genomic-editing-greatly-improves-precision/]
- 51. Engineering the Cas9 nuclease to enhance prime editing ... [https://www.nature.com/articles/s41587-025-02876-6]
- 52. Systematic optimization of prime editing for enhanced ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2025.1589034/full]